Introduction: Adopting a sedentary lifeway
5
2006
... 定居(sedentism)是指人群降低移动性,并最终在某个地方全年固定地居住下来的过程[1].定居并非一个现象,它体现了人群在移动性降低这一过程中的各种不同居住形式,如“人群全年居住于同一地点”或“至少人群中的一部分全年居住于同一地点”等[2].这些形式均体现出定居作为一种特殊的社会地理形态,其社会文化模式在一定区域范围和一段时间内固定并维持[3].区别于随机移动、周期性移动的社会特征,定居的产生是对人群过去长期存在的流动性的根本突破[4-5]. ...
... 最早的定居发生在农业出现之前的狩猎采集人群中,这一观点在学界已达成共识[1],但是具体到各地区和考古学文化,定居出现的时间及其与农业的关系存在广泛的差异[7].西亚地区最早的定居发生在纳图夫文化(Natufian Culture)时期(13000—10000 a BP),集中于黎凡特(Levant)中南部地区[8-9],比该地区农业社会的出现早了至少3000 a[10-11].中美洲地区的植物驯化始于10000—8000 a BP,但直到约4500 a BP才出现定居的农业村落[12].在东亚地区,渔猎采集经济支撑了日本绳纹时代定居村落的产生(~9800 a BP),其中晚期也出现了社会复杂化的现象[13⇓-15].中国在全新世早期的浙江浦阳上山遗址(10000—8000 a BP)已显现出定居村落的特征,其出现与水稻驯化的开始密切相关.直到现在,仍然存在以渔猎—采集为生的定居聚落,如分布于北极和亚北极地区的楚克奇人(Chukchi)、因纽特人(Inuit)和阿留申人(Aleutian)[3,16-17],也有进行狩猎采集、同时进行农业生产的定居聚落,如南非地区的布须曼人(Bushmen)[18]和亚马逊低地的印第安人[19].上述的考古学和民族学证据表明,世界范围内各地区人群定居出现时间存在差异,且定居的经济基础形式多样[20]. ...
... 而定居的产生则被归结为气候变化、人口增长、资源压力、技术革新等自然环境和社会经济因素共同作用的结果[21-22].“推拉假说”("Push" and "Pull" Hypotheses)强调丰富的自然资源或由干旱化等因素造成的环境压力以“拉”或“推”的方式促使人们向环境较好的地区聚集并定居[23].根据“拉”理论的假设,丰富的资源吸引人们减少流动性进而产生定居,比如非洲北部地区新石器时代早期,常年或季节性河流提供的丰富渔猎资源对狩猎采集者的吸引、纳图夫文化时期先民对瞪羚(Gazella gazella)等非长距迁徙动物和季节性植物资源利用,促使非农业狩猎采集社会的移动性降低、定居村落产生[24⇓-26].“推”指狩猎采集人群受到环境变化或生计压力被迫转向定居生活,一方面强调气候变化因素的影响,如干旱化导致的资源萎缩促使人们向环境较好的地区聚集并定居[27-28],另一方面强调人口增长导致的资源压力增加[2,29 -30],促使人群向定居转变.还有一种解释认为,区域性的人口增长导致各群体内部联系更加紧密,在当地资源枯竭时更难迁移从而驱动定居.随后定居在群体间扩散,进一步通过提高生育率促进人口增长,不断正反馈强化“定居—人口增长—定居”这一过程,使定居程度不断加深[13,31].“海洋假说”(Maritime Hypothesis)认为,一些以海洋资源为经济基础的社会可能出现类似农业的经济集约化,并趋于定居生活,并且也像以农业为基础的社会一样具有社会复杂性[1];另外,地理环境差异造成的资源不均匀分布会促使狩猎采集人群降低移动性,趋于定居,生态交错带等资源关键区则发挥了关键作用[7,32 -33].从经济和政治等社会因素角度来看,人口增长、食物储藏技术的发展、领地意识的产生、个人财产权的出现使社会连结更加紧密,人们主动或被动的加入定居的生活方式中,以分散个体所承担的风险、增加合作、提高资源获取效率、促进技术发展和人口增长以应对环境变化[9,13,20,22,32,34⇓⇓⇓⇓⇓⇓ -41].随后定居的生活方式在群体间扩散,随着不断正反馈强化,使定居程度不断加深[13,31]. ...
... 判断一个遗址是否出现定居有多条标准,包括投入大量人力物力建造耐用的建 筑、重型物质文化遗存如大型石磨盘、储藏设施、墓地、伴生动物如家鼠和家麻雀(Passer domesticus)的出现,动植物遗存所显示的遗址使用季节、陶器的出现和文化层厚度等[1,7,9,11,42 -43].尽管遗址往往无法符合以上全部条件,在判断遗址是否出现定居时仍需考虑多个因素. ...
... 季节性分析是指示遗址是否为全年居住的直接证据,也反映了人类利用自然资源 的策略和能力.温带地区动物的生长、繁殖和行为都受到季节变化的影响,因此遗址出土的动物遗存是进行遗址季节性分析的重要材料.判断方法包括但不限于:① 是否出现只在特定时间出现的物种,包括候鸟、洄游的鱼类、有迁徙习性的哺乳动物和昆虫的蛹等[107,128];② 一些骨骼的形成具有季节性特征,如鹿角生长—脱落的年度循环和雌性鸟类繁殖季节才会形成的髓质骨[104,108,129];③ 动物的出生季节相对固定,通过牙齿和骨骼判断哺乳动物的死亡年龄,可以计算得到死亡季节[108,130];④ 动物的一些部位终生生长并且受到季节变化的影响表现出周期性生长特征,通过观察生长纹可以得知动物的死亡季节,适合研究的材料包括贝壳、鱼类脊椎、鳃盖骨、耳石和哺乳动物牙骨质等[104];⑤通过食草类动物牙齿釉质氧同位素变化判断死亡季节[131].通过以上方法分析每个动物个体的死亡季节,可以得知一个物种的死亡季节分布;通过汇总所有物种所有个体的死亡季节,便可以得到遗址的使用季节.另外由于遗址堆积形成过程中受到多种埋藏学因素的影响,在利用动物遗存讨论获取季节时应尽量排除因自然营力进入遗址堆积的物种,选择明确因人类活动进入遗址堆积的样品,才能对遗址使用的季节性有更为全面、客观的了解[1,43,104]. ...
Hunter-gatherer economic complexity and "population pressure": A cross-cultural analysis
2
1988
... 定居(sedentism)是指人群降低移动性,并最终在某个地方全年固定地居住下来的过程[1].定居并非一个现象,它体现了人群在移动性降低这一过程中的各种不同居住形式,如“人群全年居住于同一地点”或“至少人群中的一部分全年居住于同一地点”等[2].这些形式均体现出定居作为一种特殊的社会地理形态,其社会文化模式在一定区域范围和一段时间内固定并维持[3].区别于随机移动、周期性移动的社会特征,定居的产生是对人群过去长期存在的流动性的根本突破[4-5]. ...
... 而定居的产生则被归结为气候变化、人口增长、资源压力、技术革新等自然环境和社会经济因素共同作用的结果[21-22].“推拉假说”("Push" and "Pull" Hypotheses)强调丰富的自然资源或由干旱化等因素造成的环境压力以“拉”或“推”的方式促使人们向环境较好的地区聚集并定居[23].根据“拉”理论的假设,丰富的资源吸引人们减少流动性进而产生定居,比如非洲北部地区新石器时代早期,常年或季节性河流提供的丰富渔猎资源对狩猎采集者的吸引、纳图夫文化时期先民对瞪羚(Gazella gazella)等非长距迁徙动物和季节性植物资源利用,促使非农业狩猎采集社会的移动性降低、定居村落产生[24⇓-26].“推”指狩猎采集人群受到环境变化或生计压力被迫转向定居生活,一方面强调气候变化因素的影响,如干旱化导致的资源萎缩促使人们向环境较好的地区聚集并定居[27-28],另一方面强调人口增长导致的资源压力增加[2,29 -30],促使人群向定居转变.还有一种解释认为,区域性的人口增长导致各群体内部联系更加紧密,在当地资源枯竭时更难迁移从而驱动定居.随后定居在群体间扩散,进一步通过提高生育率促进人口增长,不断正反馈强化“定居—人口增长—定居”这一过程,使定居程度不断加深[13,31].“海洋假说”(Maritime Hypothesis)认为,一些以海洋资源为经济基础的社会可能出现类似农业的经济集约化,并趋于定居生活,并且也像以农业为基础的社会一样具有社会复杂性[1];另外,地理环境差异造成的资源不均匀分布会促使狩猎采集人群降低移动性,趋于定居,生态交错带等资源关键区则发挥了关键作用[7,32 -33].从经济和政治等社会因素角度来看,人口增长、食物储藏技术的发展、领地意识的产生、个人财产权的出现使社会连结更加紧密,人们主动或被动的加入定居的生活方式中,以分散个体所承担的风险、增加合作、提高资源获取效率、促进技术发展和人口增长以应对环境变化[9,13,20,22,32,34⇓⇓⇓⇓⇓⇓ -41].随后定居的生活方式在群体间扩散,随着不断正反馈强化,使定居程度不断加深[13,31]. ...
Mobility/sedentism: Concepts, archaeological measures, and effects
2
1992
... 定居(sedentism)是指人群降低移动性,并最终在某个地方全年固定地居住下来的过程[1].定居并非一个现象,它体现了人群在移动性降低这一过程中的各种不同居住形式,如“人群全年居住于同一地点”或“至少人群中的一部分全年居住于同一地点”等[2].这些形式均体现出定居作为一种特殊的社会地理形态,其社会文化模式在一定区域范围和一段时间内固定并维持[3].区别于随机移动、周期性移动的社会特征,定居的产生是对人群过去长期存在的流动性的根本突破[4-5]. ...
... 最早的定居发生在农业出现之前的狩猎采集人群中,这一观点在学界已达成共识[1],但是具体到各地区和考古学文化,定居出现的时间及其与农业的关系存在广泛的差异[7].西亚地区最早的定居发生在纳图夫文化(Natufian Culture)时期(13000—10000 a BP),集中于黎凡特(Levant)中南部地区[8-9],比该地区农业社会的出现早了至少3000 a[10-11].中美洲地区的植物驯化始于10000—8000 a BP,但直到约4500 a BP才出现定居的农业村落[12].在东亚地区,渔猎采集经济支撑了日本绳纹时代定居村落的产生(~9800 a BP),其中晚期也出现了社会复杂化的现象[13⇓-15].中国在全新世早期的浙江浦阳上山遗址(10000—8000 a BP)已显现出定居村落的特征,其出现与水稻驯化的开始密切相关.直到现在,仍然存在以渔猎—采集为生的定居聚落,如分布于北极和亚北极地区的楚克奇人(Chukchi)、因纽特人(Inuit)和阿留申人(Aleutian)[3,16-17],也有进行狩猎采集、同时进行农业生产的定居聚落,如南非地区的布须曼人(Bushmen)[18]和亚马逊低地的印第安人[19].上述的考古学和民族学证据表明,世界范围内各地区人群定居出现时间存在差异,且定居的经济基础形式多样[20]. ...
2
2005
... 定居(sedentism)是指人群降低移动性,并最终在某个地方全年固定地居住下来的过程[1].定居并非一个现象,它体现了人群在移动性降低这一过程中的各种不同居住形式,如“人群全年居住于同一地点”或“至少人群中的一部分全年居住于同一地点”等[2].这些形式均体现出定居作为一种特殊的社会地理形态,其社会文化模式在一定区域范围和一段时间内固定并维持[3].区别于随机移动、周期性移动的社会特征,定居的产生是对人群过去长期存在的流动性的根本突破[4-5]. ...
... 人群从移动到定居的变化,对人类社会产生了深远的影响.定居的生活方式使得生育率提高、生育周期缩短和死亡率降低,进而促进人口指数级增长[6].同时,通过对固定区域内资源的集中开发,相同单位土地可以产出更多的能量,提高了环境承载力,从而能够养活更多的人口,甚至产生食物剩余.这些盈余的食物养活了一批从事专门活动的手工业者,从而促进了技术的发展;通过供养全职的贵族、官僚、士兵和神职人员,导致了社会分层和政治集权,使社会组织产生巨大变化[4-5]. ...
Evolution, consequences and future of plant and animal domestication
2
2002
... 定居(sedentism)是指人群降低移动性,并最终在某个地方全年固定地居住下来的过程[1].定居并非一个现象,它体现了人群在移动性降低这一过程中的各种不同居住形式,如“人群全年居住于同一地点”或“至少人群中的一部分全年居住于同一地点”等[2].这些形式均体现出定居作为一种特殊的社会地理形态,其社会文化模式在一定区域范围和一段时间内固定并维持[3].区别于随机移动、周期性移动的社会特征,定居的产生是对人群过去长期存在的流动性的根本突破[4-5]. ...
... 人群从移动到定居的变化,对人类社会产生了深远的影响.定居的生活方式使得生育率提高、生育周期缩短和死亡率降低,进而促进人口指数级增长[6].同时,通过对固定区域内资源的集中开发,相同单位土地可以产出更多的能量,提高了环境承载力,从而能够养活更多的人口,甚至产生食物剩余.这些盈余的食物养活了一批从事专门活动的手工业者,从而促进了技术的发展;通过供养全职的贵族、官僚、士兵和神职人员,导致了社会分层和政治集权,使社会组织产生巨大变化[4-5]. ...
From sedentism to states, 10 000-3000 BCE//Daniel C S. A Companion to the Ancient Near East. 2nd ed
1
2020
... 人群从移动到定居的变化,对人类社会产生了深远的影响.定居的生活方式使得生育率提高、生育周期缩短和死亡率降低,进而促进人口指数级增长[6].同时,通过对固定区域内资源的集中开发,相同单位土地可以产出更多的能量,提高了环境承载力,从而能够养活更多的人口,甚至产生食物剩余.这些盈余的食物养活了一批从事专门活动的手工业者,从而促进了技术的发展;通过供养全职的贵族、官僚、士兵和神职人员,导致了社会分层和政治集权,使社会组织产生巨大变化[4-5]. ...
3
2000
... 最早的定居发生在农业出现之前的狩猎采集人群中,这一观点在学界已达成共识[1],但是具体到各地区和考古学文化,定居出现的时间及其与农业的关系存在广泛的差异[7].西亚地区最早的定居发生在纳图夫文化(Natufian Culture)时期(13000—10000 a BP),集中于黎凡特(Levant)中南部地区[8-9],比该地区农业社会的出现早了至少3000 a[10-11].中美洲地区的植物驯化始于10000—8000 a BP,但直到约4500 a BP才出现定居的农业村落[12].在东亚地区,渔猎采集经济支撑了日本绳纹时代定居村落的产生(~9800 a BP),其中晚期也出现了社会复杂化的现象[13⇓-15].中国在全新世早期的浙江浦阳上山遗址(10000—8000 a BP)已显现出定居村落的特征,其出现与水稻驯化的开始密切相关.直到现在,仍然存在以渔猎—采集为生的定居聚落,如分布于北极和亚北极地区的楚克奇人(Chukchi)、因纽特人(Inuit)和阿留申人(Aleutian)[3,16-17],也有进行狩猎采集、同时进行农业生产的定居聚落,如南非地区的布须曼人(Bushmen)[18]和亚马逊低地的印第安人[19].上述的考古学和民族学证据表明,世界范围内各地区人群定居出现时间存在差异,且定居的经济基础形式多样[20]. ...
... 而定居的产生则被归结为气候变化、人口增长、资源压力、技术革新等自然环境和社会经济因素共同作用的结果[21-22].“推拉假说”("Push" and "Pull" Hypotheses)强调丰富的自然资源或由干旱化等因素造成的环境压力以“拉”或“推”的方式促使人们向环境较好的地区聚集并定居[23].根据“拉”理论的假设,丰富的资源吸引人们减少流动性进而产生定居,比如非洲北部地区新石器时代早期,常年或季节性河流提供的丰富渔猎资源对狩猎采集者的吸引、纳图夫文化时期先民对瞪羚(Gazella gazella)等非长距迁徙动物和季节性植物资源利用,促使非农业狩猎采集社会的移动性降低、定居村落产生[24⇓-26].“推”指狩猎采集人群受到环境变化或生计压力被迫转向定居生活,一方面强调气候变化因素的影响,如干旱化导致的资源萎缩促使人们向环境较好的地区聚集并定居[27-28],另一方面强调人口增长导致的资源压力增加[2,29 -30],促使人群向定居转变.还有一种解释认为,区域性的人口增长导致各群体内部联系更加紧密,在当地资源枯竭时更难迁移从而驱动定居.随后定居在群体间扩散,进一步通过提高生育率促进人口增长,不断正反馈强化“定居—人口增长—定居”这一过程,使定居程度不断加深[13,31].“海洋假说”(Maritime Hypothesis)认为,一些以海洋资源为经济基础的社会可能出现类似农业的经济集约化,并趋于定居生活,并且也像以农业为基础的社会一样具有社会复杂性[1];另外,地理环境差异造成的资源不均匀分布会促使狩猎采集人群降低移动性,趋于定居,生态交错带等资源关键区则发挥了关键作用[7,32 -33].从经济和政治等社会因素角度来看,人口增长、食物储藏技术的发展、领地意识的产生、个人财产权的出现使社会连结更加紧密,人们主动或被动的加入定居的生活方式中,以分散个体所承担的风险、增加合作、提高资源获取效率、促进技术发展和人口增长以应对环境变化[9,13,20,22,32,34⇓⇓⇓⇓⇓⇓ -41].随后定居的生活方式在群体间扩散,随着不断正反馈强化,使定居程度不断加深[13,31]. ...
... 判断一个遗址是否出现定居有多条标准,包括投入大量人力物力建造耐用的建 筑、重型物质文化遗存如大型石磨盘、储藏设施、墓地、伴生动物如家鼠和家麻雀(Passer domesticus)的出现,动植物遗存所显示的遗址使用季节、陶器的出现和文化层厚度等[1,7,9,11,42 -43].尽管遗址往往无法符合以上全部条件,在判断遗址是否出现定居时仍需考虑多个因素. ...
Foraging, farming, and social complexity in the pre-pottery Neolithic of the southern Levant: A review and synthesis
1
2002
... 最早的定居发生在农业出现之前的狩猎采集人群中,这一观点在学界已达成共识[1],但是具体到各地区和考古学文化,定居出现的时间及其与农业的关系存在广泛的差异[7].西亚地区最早的定居发生在纳图夫文化(Natufian Culture)时期(13000—10000 a BP),集中于黎凡特(Levant)中南部地区[8-9],比该地区农业社会的出现早了至少3000 a[10-11].中美洲地区的植物驯化始于10000—8000 a BP,但直到约4500 a BP才出现定居的农业村落[12].在东亚地区,渔猎采集经济支撑了日本绳纹时代定居村落的产生(~9800 a BP),其中晚期也出现了社会复杂化的现象[13⇓-15].中国在全新世早期的浙江浦阳上山遗址(10000—8000 a BP)已显现出定居村落的特征,其出现与水稻驯化的开始密切相关.直到现在,仍然存在以渔猎—采集为生的定居聚落,如分布于北极和亚北极地区的楚克奇人(Chukchi)、因纽特人(Inuit)和阿留申人(Aleutian)[3,16-17],也有进行狩猎采集、同时进行农业生产的定居聚落,如南非地区的布须曼人(Bushmen)[18]和亚马逊低地的印第安人[19].上述的考古学和民族学证据表明,世界范围内各地区人群定居出现时间存在差异,且定居的经济基础形式多样[20]. ...
Preagricultural sedentism:The Natufian example//Price T D, Brown J A
3
1985
... 最早的定居发生在农业出现之前的狩猎采集人群中,这一观点在学界已达成共识[1],但是具体到各地区和考古学文化,定居出现的时间及其与农业的关系存在广泛的差异[7].西亚地区最早的定居发生在纳图夫文化(Natufian Culture)时期(13000—10000 a BP),集中于黎凡特(Levant)中南部地区[8-9],比该地区农业社会的出现早了至少3000 a[10-11].中美洲地区的植物驯化始于10000—8000 a BP,但直到约4500 a BP才出现定居的农业村落[12].在东亚地区,渔猎采集经济支撑了日本绳纹时代定居村落的产生(~9800 a BP),其中晚期也出现了社会复杂化的现象[13⇓-15].中国在全新世早期的浙江浦阳上山遗址(10000—8000 a BP)已显现出定居村落的特征,其出现与水稻驯化的开始密切相关.直到现在,仍然存在以渔猎—采集为生的定居聚落,如分布于北极和亚北极地区的楚克奇人(Chukchi)、因纽特人(Inuit)和阿留申人(Aleutian)[3,16-17],也有进行狩猎采集、同时进行农业生产的定居聚落,如南非地区的布须曼人(Bushmen)[18]和亚马逊低地的印第安人[19].上述的考古学和民族学证据表明,世界范围内各地区人群定居出现时间存在差异,且定居的经济基础形式多样[20]. ...
... 而定居的产生则被归结为气候变化、人口增长、资源压力、技术革新等自然环境和社会经济因素共同作用的结果[21-22].“推拉假说”("Push" and "Pull" Hypotheses)强调丰富的自然资源或由干旱化等因素造成的环境压力以“拉”或“推”的方式促使人们向环境较好的地区聚集并定居[23].根据“拉”理论的假设,丰富的资源吸引人们减少流动性进而产生定居,比如非洲北部地区新石器时代早期,常年或季节性河流提供的丰富渔猎资源对狩猎采集者的吸引、纳图夫文化时期先民对瞪羚(Gazella gazella)等非长距迁徙动物和季节性植物资源利用,促使非农业狩猎采集社会的移动性降低、定居村落产生[24⇓-26].“推”指狩猎采集人群受到环境变化或生计压力被迫转向定居生活,一方面强调气候变化因素的影响,如干旱化导致的资源萎缩促使人们向环境较好的地区聚集并定居[27-28],另一方面强调人口增长导致的资源压力增加[2,29 -30],促使人群向定居转变.还有一种解释认为,区域性的人口增长导致各群体内部联系更加紧密,在当地资源枯竭时更难迁移从而驱动定居.随后定居在群体间扩散,进一步通过提高生育率促进人口增长,不断正反馈强化“定居—人口增长—定居”这一过程,使定居程度不断加深[13,31].“海洋假说”(Maritime Hypothesis)认为,一些以海洋资源为经济基础的社会可能出现类似农业的经济集约化,并趋于定居生活,并且也像以农业为基础的社会一样具有社会复杂性[1];另外,地理环境差异造成的资源不均匀分布会促使狩猎采集人群降低移动性,趋于定居,生态交错带等资源关键区则发挥了关键作用[7,32 -33].从经济和政治等社会因素角度来看,人口增长、食物储藏技术的发展、领地意识的产生、个人财产权的出现使社会连结更加紧密,人们主动或被动的加入定居的生活方式中,以分散个体所承担的风险、增加合作、提高资源获取效率、促进技术发展和人口增长以应对环境变化[9,13,20,22,32,34⇓⇓⇓⇓⇓⇓ -41].随后定居的生活方式在群体间扩散,随着不断正反馈强化,使定居程度不断加深[13,31]. ...
... 判断一个遗址是否出现定居有多条标准,包括投入大量人力物力建造耐用的建 筑、重型物质文化遗存如大型石磨盘、储藏设施、墓地、伴生动物如家鼠和家麻雀(Passer domesticus)的出现,动植物遗存所显示的遗址使用季节、陶器的出现和文化层厚度等[1,7,9,11,42 -43].尽管遗址往往无法符合以上全部条件,在判断遗址是否出现定居时仍需考虑多个因素. ...
From Pleistocene to Holocene: The prehistory of southwest Asia in evolutionary context
1
2017
... 最早的定居发生在农业出现之前的狩猎采集人群中,这一观点在学界已达成共识[1],但是具体到各地区和考古学文化,定居出现的时间及其与农业的关系存在广泛的差异[7].西亚地区最早的定居发生在纳图夫文化(Natufian Culture)时期(13000—10000 a BP),集中于黎凡特(Levant)中南部地区[8-9],比该地区农业社会的出现早了至少3000 a[10-11].中美洲地区的植物驯化始于10000—8000 a BP,但直到约4500 a BP才出现定居的农业村落[12].在东亚地区,渔猎采集经济支撑了日本绳纹时代定居村落的产生(~9800 a BP),其中晚期也出现了社会复杂化的现象[13⇓-15].中国在全新世早期的浙江浦阳上山遗址(10000—8000 a BP)已显现出定居村落的特征,其出现与水稻驯化的开始密切相关.直到现在,仍然存在以渔猎—采集为生的定居聚落,如分布于北极和亚北极地区的楚克奇人(Chukchi)、因纽特人(Inuit)和阿留申人(Aleutian)[3,16-17],也有进行狩猎采集、同时进行农业生产的定居聚落,如南非地区的布须曼人(Bushmen)[18]和亚马逊低地的印第安人[19].上述的考古学和民族学证据表明,世界范围内各地区人群定居出现时间存在差异,且定居的经济基础形式多样[20]. ...
On "sedentism" in the Later Epipalaeolithic (Natufian) Levant
2
2006
... 最早的定居发生在农业出现之前的狩猎采集人群中,这一观点在学界已达成共识[1],但是具体到各地区和考古学文化,定居出现的时间及其与农业的关系存在广泛的差异[7].西亚地区最早的定居发生在纳图夫文化(Natufian Culture)时期(13000—10000 a BP),集中于黎凡特(Levant)中南部地区[8-9],比该地区农业社会的出现早了至少3000 a[10-11].中美洲地区的植物驯化始于10000—8000 a BP,但直到约4500 a BP才出现定居的农业村落[12].在东亚地区,渔猎采集经济支撑了日本绳纹时代定居村落的产生(~9800 a BP),其中晚期也出现了社会复杂化的现象[13⇓-15].中国在全新世早期的浙江浦阳上山遗址(10000—8000 a BP)已显现出定居村落的特征,其出现与水稻驯化的开始密切相关.直到现在,仍然存在以渔猎—采集为生的定居聚落,如分布于北极和亚北极地区的楚克奇人(Chukchi)、因纽特人(Inuit)和阿留申人(Aleutian)[3,16-17],也有进行狩猎采集、同时进行农业生产的定居聚落,如南非地区的布须曼人(Bushmen)[18]和亚马逊低地的印第安人[19].上述的考古学和民族学证据表明,世界范围内各地区人群定居出现时间存在差异,且定居的经济基础形式多样[20]. ...
... 判断一个遗址是否出现定居有多条标准,包括投入大量人力物力建造耐用的建 筑、重型物质文化遗存如大型石磨盘、储藏设施、墓地、伴生动物如家鼠和家麻雀(Passer domesticus)的出现,动植物遗存所显示的遗址使用季节、陶器的出现和文化层厚度等[1,7,9,11,42 -43].尽管遗址往往无法符合以上全部条件,在判断遗址是否出现定居时仍需考虑多个因素. ...
Low-level food production
1
2001
... 最早的定居发生在农业出现之前的狩猎采集人群中,这一观点在学界已达成共识[1],但是具体到各地区和考古学文化,定居出现的时间及其与农业的关系存在广泛的差异[7].西亚地区最早的定居发生在纳图夫文化(Natufian Culture)时期(13000—10000 a BP),集中于黎凡特(Levant)中南部地区[8-9],比该地区农业社会的出现早了至少3000 a[10-11].中美洲地区的植物驯化始于10000—8000 a BP,但直到约4500 a BP才出现定居的农业村落[12].在东亚地区,渔猎采集经济支撑了日本绳纹时代定居村落的产生(~9800 a BP),其中晚期也出现了社会复杂化的现象[13⇓-15].中国在全新世早期的浙江浦阳上山遗址(10000—8000 a BP)已显现出定居村落的特征,其出现与水稻驯化的开始密切相关.直到现在,仍然存在以渔猎—采集为生的定居聚落,如分布于北极和亚北极地区的楚克奇人(Chukchi)、因纽特人(Inuit)和阿留申人(Aleutian)[3,16-17],也有进行狩猎采集、同时进行农业生产的定居聚落,如南非地区的布须曼人(Bushmen)[18]和亚马逊低地的印第安人[19].上述的考古学和民族学证据表明,世界范围内各地区人群定居出现时间存在差异,且定居的经济基础形式多样[20]. ...
Jomon hot spot: Increasing sedentism in south-western Japan in the Incipient Jomon (14000-9250 cal. BC) and Earliest Jomon (9250-5300 cal. BC) periods
4
2006
... 最早的定居发生在农业出现之前的狩猎采集人群中,这一观点在学界已达成共识[1],但是具体到各地区和考古学文化,定居出现的时间及其与农业的关系存在广泛的差异[7].西亚地区最早的定居发生在纳图夫文化(Natufian Culture)时期(13000—10000 a BP),集中于黎凡特(Levant)中南部地区[8-9],比该地区农业社会的出现早了至少3000 a[10-11].中美洲地区的植物驯化始于10000—8000 a BP,但直到约4500 a BP才出现定居的农业村落[12].在东亚地区,渔猎采集经济支撑了日本绳纹时代定居村落的产生(~9800 a BP),其中晚期也出现了社会复杂化的现象[13⇓-15].中国在全新世早期的浙江浦阳上山遗址(10000—8000 a BP)已显现出定居村落的特征,其出现与水稻驯化的开始密切相关.直到现在,仍然存在以渔猎—采集为生的定居聚落,如分布于北极和亚北极地区的楚克奇人(Chukchi)、因纽特人(Inuit)和阿留申人(Aleutian)[3,16-17],也有进行狩猎采集、同时进行农业生产的定居聚落,如南非地区的布须曼人(Bushmen)[18]和亚马逊低地的印第安人[19].上述的考古学和民族学证据表明,世界范围内各地区人群定居出现时间存在差异,且定居的经济基础形式多样[20]. ...
... 而定居的产生则被归结为气候变化、人口增长、资源压力、技术革新等自然环境和社会经济因素共同作用的结果[21-22].“推拉假说”("Push" and "Pull" Hypotheses)强调丰富的自然资源或由干旱化等因素造成的环境压力以“拉”或“推”的方式促使人们向环境较好的地区聚集并定居[23].根据“拉”理论的假设,丰富的资源吸引人们减少流动性进而产生定居,比如非洲北部地区新石器时代早期,常年或季节性河流提供的丰富渔猎资源对狩猎采集者的吸引、纳图夫文化时期先民对瞪羚(Gazella gazella)等非长距迁徙动物和季节性植物资源利用,促使非农业狩猎采集社会的移动性降低、定居村落产生[24⇓-26].“推”指狩猎采集人群受到环境变化或生计压力被迫转向定居生活,一方面强调气候变化因素的影响,如干旱化导致的资源萎缩促使人们向环境较好的地区聚集并定居[27-28],另一方面强调人口增长导致的资源压力增加[2,29 -30],促使人群向定居转变.还有一种解释认为,区域性的人口增长导致各群体内部联系更加紧密,在当地资源枯竭时更难迁移从而驱动定居.随后定居在群体间扩散,进一步通过提高生育率促进人口增长,不断正反馈强化“定居—人口增长—定居”这一过程,使定居程度不断加深[13,31].“海洋假说”(Maritime Hypothesis)认为,一些以海洋资源为经济基础的社会可能出现类似农业的经济集约化,并趋于定居生活,并且也像以农业为基础的社会一样具有社会复杂性[1];另外,地理环境差异造成的资源不均匀分布会促使狩猎采集人群降低移动性,趋于定居,生态交错带等资源关键区则发挥了关键作用[7,32 -33].从经济和政治等社会因素角度来看,人口增长、食物储藏技术的发展、领地意识的产生、个人财产权的出现使社会连结更加紧密,人们主动或被动的加入定居的生活方式中,以分散个体所承担的风险、增加合作、提高资源获取效率、促进技术发展和人口增长以应对环境变化[9,13,20,22,32,34⇓⇓⇓⇓⇓⇓ -41].随后定居的生活方式在群体间扩散,随着不断正反馈强化,使定居程度不断加深[13,31]. ...
... ,13,20,22,32,34⇓⇓⇓⇓⇓⇓ -41].随后定居的生活方式在群体间扩散,随着不断正反馈强化,使定居程度不断加深[13,31]. ...
... [13,31]. ...
1
2004
... 最早的定居发生在农业出现之前的狩猎采集人群中,这一观点在学界已达成共识[1],但是具体到各地区和考古学文化,定居出现的时间及其与农业的关系存在广泛的差异[7].西亚地区最早的定居发生在纳图夫文化(Natufian Culture)时期(13000—10000 a BP),集中于黎凡特(Levant)中南部地区[8-9],比该地区农业社会的出现早了至少3000 a[10-11].中美洲地区的植物驯化始于10000—8000 a BP,但直到约4500 a BP才出现定居的农业村落[12].在东亚地区,渔猎采集经济支撑了日本绳纹时代定居村落的产生(~9800 a BP),其中晚期也出现了社会复杂化的现象[13⇓-15].中国在全新世早期的浙江浦阳上山遗址(10000—8000 a BP)已显现出定居村落的特征,其出现与水稻驯化的开始密切相关.直到现在,仍然存在以渔猎—采集为生的定居聚落,如分布于北极和亚北极地区的楚克奇人(Chukchi)、因纽特人(Inuit)和阿留申人(Aleutian)[3,16-17],也有进行狩猎采集、同时进行农业生产的定居聚落,如南非地区的布须曼人(Bushmen)[18]和亚马逊低地的印第安人[19].上述的考古学和民族学证据表明,世界范围内各地区人群定居出现时间存在差异,且定居的经济基础形式多样[20]. ...
Growth and decline in complex hunter-gatherer societies: A case study from the Jomon period Sannai Maruyama site
1
2008
... 最早的定居发生在农业出现之前的狩猎采集人群中,这一观点在学界已达成共识[1],但是具体到各地区和考古学文化,定居出现的时间及其与农业的关系存在广泛的差异[7].西亚地区最早的定居发生在纳图夫文化(Natufian Culture)时期(13000—10000 a BP),集中于黎凡特(Levant)中南部地区[8-9],比该地区农业社会的出现早了至少3000 a[10-11].中美洲地区的植物驯化始于10000—8000 a BP,但直到约4500 a BP才出现定居的农业村落[12].在东亚地区,渔猎采集经济支撑了日本绳纹时代定居村落的产生(~9800 a BP),其中晚期也出现了社会复杂化的现象[13⇓-15].中国在全新世早期的浙江浦阳上山遗址(10000—8000 a BP)已显现出定居村落的特征,其出现与水稻驯化的开始密切相关.直到现在,仍然存在以渔猎—采集为生的定居聚落,如分布于北极和亚北极地区的楚克奇人(Chukchi)、因纽特人(Inuit)和阿留申人(Aleutian)[3,16-17],也有进行狩猎采集、同时进行农业生产的定居聚落,如南非地区的布须曼人(Bushmen)[18]和亚马逊低地的印第安人[19].上述的考古学和民族学证据表明,世界范围内各地区人群定居出现时间存在差异,且定居的经济基础形式多样[20]. ...
Maintaining cohesion through rituals: Chukchi herders and hunters, a people of the Siberian Arctic
1
2005
... 最早的定居发生在农业出现之前的狩猎采集人群中,这一观点在学界已达成共识[1],但是具体到各地区和考古学文化,定居出现的时间及其与农业的关系存在广泛的差异[7].西亚地区最早的定居发生在纳图夫文化(Natufian Culture)时期(13000—10000 a BP),集中于黎凡特(Levant)中南部地区[8-9],比该地区农业社会的出现早了至少3000 a[10-11].中美洲地区的植物驯化始于10000—8000 a BP,但直到约4500 a BP才出现定居的农业村落[12].在东亚地区,渔猎采集经济支撑了日本绳纹时代定居村落的产生(~9800 a BP),其中晚期也出现了社会复杂化的现象[13⇓-15].中国在全新世早期的浙江浦阳上山遗址(10000—8000 a BP)已显现出定居村落的特征,其出现与水稻驯化的开始密切相关.直到现在,仍然存在以渔猎—采集为生的定居聚落,如分布于北极和亚北极地区的楚克奇人(Chukchi)、因纽特人(Inuit)和阿留申人(Aleutian)[3,16-17],也有进行狩猎采集、同时进行农业生产的定居聚落,如南非地区的布须曼人(Bushmen)[18]和亚马逊低地的印第安人[19].上述的考古学和民族学证据表明,世界范围内各地区人群定居出现时间存在差异,且定居的经济基础形式多样[20]. ...
The evolution of complex hunter-gatherers on the Kodiak Archipelago
1
2003
... 最早的定居发生在农业出现之前的狩猎采集人群中,这一观点在学界已达成共识[1],但是具体到各地区和考古学文化,定居出现的时间及其与农业的关系存在广泛的差异[7].西亚地区最早的定居发生在纳图夫文化(Natufian Culture)时期(13000—10000 a BP),集中于黎凡特(Levant)中南部地区[8-9],比该地区农业社会的出现早了至少3000 a[10-11].中美洲地区的植物驯化始于10000—8000 a BP,但直到约4500 a BP才出现定居的农业村落[12].在东亚地区,渔猎采集经济支撑了日本绳纹时代定居村落的产生(~9800 a BP),其中晚期也出现了社会复杂化的现象[13⇓-15].中国在全新世早期的浙江浦阳上山遗址(10000—8000 a BP)已显现出定居村落的特征,其出现与水稻驯化的开始密切相关.直到现在,仍然存在以渔猎—采集为生的定居聚落,如分布于北极和亚北极地区的楚克奇人(Chukchi)、因纽特人(Inuit)和阿留申人(Aleutian)[3,16-17],也有进行狩猎采集、同时进行农业生产的定居聚落,如南非地区的布须曼人(Bushmen)[18]和亚马逊低地的印第安人[19].上述的考古学和民族学证据表明,世界范围内各地区人群定居出现时间存在差异,且定居的经济基础形式多样[20]. ...
1
1972
... 最早的定居发生在农业出现之前的狩猎采集人群中,这一观点在学界已达成共识[1],但是具体到各地区和考古学文化,定居出现的时间及其与农业的关系存在广泛的差异[7].西亚地区最早的定居发生在纳图夫文化(Natufian Culture)时期(13000—10000 a BP),集中于黎凡特(Levant)中南部地区[8-9],比该地区农业社会的出现早了至少3000 a[10-11].中美洲地区的植物驯化始于10000—8000 a BP,但直到约4500 a BP才出现定居的农业村落[12].在东亚地区,渔猎采集经济支撑了日本绳纹时代定居村落的产生(~9800 a BP),其中晚期也出现了社会复杂化的现象[13⇓-15].中国在全新世早期的浙江浦阳上山遗址(10000—8000 a BP)已显现出定居村落的特征,其出现与水稻驯化的开始密切相关.直到现在,仍然存在以渔猎—采集为生的定居聚落,如分布于北极和亚北极地区的楚克奇人(Chukchi)、因纽特人(Inuit)和阿留申人(Aleutian)[3,16-17],也有进行狩猎采集、同时进行农业生产的定居聚落,如南非地区的布须曼人(Bushmen)[18]和亚马逊低地的印第安人[19].上述的考古学和民族学证据表明,世界范围内各地区人群定居出现时间存在差异,且定居的经济基础形式多样[20]. ...
Trekking through history: The huaorani of Amazonian Ecuador
1
2004
... 最早的定居发生在农业出现之前的狩猎采集人群中,这一观点在学界已达成共识[1],但是具体到各地区和考古学文化,定居出现的时间及其与农业的关系存在广泛的差异[7].西亚地区最早的定居发生在纳图夫文化(Natufian Culture)时期(13000—10000 a BP),集中于黎凡特(Levant)中南部地区[8-9],比该地区农业社会的出现早了至少3000 a[10-11].中美洲地区的植物驯化始于10000—8000 a BP,但直到约4500 a BP才出现定居的农业村落[12].在东亚地区,渔猎采集经济支撑了日本绳纹时代定居村落的产生(~9800 a BP),其中晚期也出现了社会复杂化的现象[13⇓-15].中国在全新世早期的浙江浦阳上山遗址(10000—8000 a BP)已显现出定居村落的特征,其出现与水稻驯化的开始密切相关.直到现在,仍然存在以渔猎—采集为生的定居聚落,如分布于北极和亚北极地区的楚克奇人(Chukchi)、因纽特人(Inuit)和阿留申人(Aleutian)[3,16-17],也有进行狩猎采集、同时进行农业生产的定居聚落,如南非地区的布须曼人(Bushmen)[18]和亚马逊低地的印第安人[19].上述的考古学和民族学证据表明,世界范围内各地区人群定居出现时间存在差异,且定居的经济基础形式多样[20]. ...
2
1983
... 最早的定居发生在农业出现之前的狩猎采集人群中,这一观点在学界已达成共识[1],但是具体到各地区和考古学文化,定居出现的时间及其与农业的关系存在广泛的差异[7].西亚地区最早的定居发生在纳图夫文化(Natufian Culture)时期(13000—10000 a BP),集中于黎凡特(Levant)中南部地区[8-9],比该地区农业社会的出现早了至少3000 a[10-11].中美洲地区的植物驯化始于10000—8000 a BP,但直到约4500 a BP才出现定居的农业村落[12].在东亚地区,渔猎采集经济支撑了日本绳纹时代定居村落的产生(~9800 a BP),其中晚期也出现了社会复杂化的现象[13⇓-15].中国在全新世早期的浙江浦阳上山遗址(10000—8000 a BP)已显现出定居村落的特征,其出现与水稻驯化的开始密切相关.直到现在,仍然存在以渔猎—采集为生的定居聚落,如分布于北极和亚北极地区的楚克奇人(Chukchi)、因纽特人(Inuit)和阿留申人(Aleutian)[3,16-17],也有进行狩猎采集、同时进行农业生产的定居聚落,如南非地区的布须曼人(Bushmen)[18]和亚马逊低地的印第安人[19].上述的考古学和民族学证据表明,世界范围内各地区人群定居出现时间存在差异,且定居的经济基础形式多样[20]. ...
... 而定居的产生则被归结为气候变化、人口增长、资源压力、技术革新等自然环境和社会经济因素共同作用的结果[21-22].“推拉假说”("Push" and "Pull" Hypotheses)强调丰富的自然资源或由干旱化等因素造成的环境压力以“拉”或“推”的方式促使人们向环境较好的地区聚集并定居[23].根据“拉”理论的假设,丰富的资源吸引人们减少流动性进而产生定居,比如非洲北部地区新石器时代早期,常年或季节性河流提供的丰富渔猎资源对狩猎采集者的吸引、纳图夫文化时期先民对瞪羚(Gazella gazella)等非长距迁徙动物和季节性植物资源利用,促使非农业狩猎采集社会的移动性降低、定居村落产生[24⇓-26].“推”指狩猎采集人群受到环境变化或生计压力被迫转向定居生活,一方面强调气候变化因素的影响,如干旱化导致的资源萎缩促使人们向环境较好的地区聚集并定居[27-28],另一方面强调人口增长导致的资源压力增加[2,29 -30],促使人群向定居转变.还有一种解释认为,区域性的人口增长导致各群体内部联系更加紧密,在当地资源枯竭时更难迁移从而驱动定居.随后定居在群体间扩散,进一步通过提高生育率促进人口增长,不断正反馈强化“定居—人口增长—定居”这一过程,使定居程度不断加深[13,31].“海洋假说”(Maritime Hypothesis)认为,一些以海洋资源为经济基础的社会可能出现类似农业的经济集约化,并趋于定居生活,并且也像以农业为基础的社会一样具有社会复杂性[1];另外,地理环境差异造成的资源不均匀分布会促使狩猎采集人群降低移动性,趋于定居,生态交错带等资源关键区则发挥了关键作用[7,32 -33].从经济和政治等社会因素角度来看,人口增长、食物储藏技术的发展、领地意识的产生、个人财产权的出现使社会连结更加紧密,人们主动或被动的加入定居的生活方式中,以分散个体所承担的风险、增加合作、提高资源获取效率、促进技术发展和人口增长以应对环境变化[9,13,20,22,32,34⇓⇓⇓⇓⇓⇓ -41].随后定居的生活方式在群体间扩散,随着不断正反馈强化,使定居程度不断加深[13,31]. ...
The origins of sedentism: Climate, population, and technology
1
2015
... 而定居的产生则被归结为气候变化、人口增长、资源压力、技术革新等自然环境和社会经济因素共同作用的结果[21-22].“推拉假说”("Push" and "Pull" Hypotheses)强调丰富的自然资源或由干旱化等因素造成的环境压力以“拉”或“推”的方式促使人们向环境较好的地区聚集并定居[23].根据“拉”理论的假设,丰富的资源吸引人们减少流动性进而产生定居,比如非洲北部地区新石器时代早期,常年或季节性河流提供的丰富渔猎资源对狩猎采集者的吸引、纳图夫文化时期先民对瞪羚(Gazella gazella)等非长距迁徙动物和季节性植物资源利用,促使非农业狩猎采集社会的移动性降低、定居村落产生[24⇓-26].“推”指狩猎采集人群受到环境变化或生计压力被迫转向定居生活,一方面强调气候变化因素的影响,如干旱化导致的资源萎缩促使人们向环境较好的地区聚集并定居[27-28],另一方面强调人口增长导致的资源压力增加[2,29 -30],促使人群向定居转变.还有一种解释认为,区域性的人口增长导致各群体内部联系更加紧密,在当地资源枯竭时更难迁移从而驱动定居.随后定居在群体间扩散,进一步通过提高生育率促进人口增长,不断正反馈强化“定居—人口增长—定居”这一过程,使定居程度不断加深[13,31].“海洋假说”(Maritime Hypothesis)认为,一些以海洋资源为经济基础的社会可能出现类似农业的经济集约化,并趋于定居生活,并且也像以农业为基础的社会一样具有社会复杂性[1];另外,地理环境差异造成的资源不均匀分布会促使狩猎采集人群降低移动性,趋于定居,生态交错带等资源关键区则发挥了关键作用[7,32 -33].从经济和政治等社会因素角度来看,人口增长、食物储藏技术的发展、领地意识的产生、个人财产权的出现使社会连结更加紧密,人们主动或被动的加入定居的生活方式中,以分散个体所承担的风险、增加合作、提高资源获取效率、促进技术发展和人口增长以应对环境变化[9,13,20,22,32,34⇓⇓⇓⇓⇓⇓ -41].随后定居的生活方式在群体间扩散,随着不断正反馈强化,使定居程度不断加深[13,31]. ...
Coevolution of farming and private property during the early Holocene
2
2013
... 而定居的产生则被归结为气候变化、人口增长、资源压力、技术革新等自然环境和社会经济因素共同作用的结果[21-22].“推拉假说”("Push" and "Pull" Hypotheses)强调丰富的自然资源或由干旱化等因素造成的环境压力以“拉”或“推”的方式促使人们向环境较好的地区聚集并定居[23].根据“拉”理论的假设,丰富的资源吸引人们减少流动性进而产生定居,比如非洲北部地区新石器时代早期,常年或季节性河流提供的丰富渔猎资源对狩猎采集者的吸引、纳图夫文化时期先民对瞪羚(Gazella gazella)等非长距迁徙动物和季节性植物资源利用,促使非农业狩猎采集社会的移动性降低、定居村落产生[24⇓-26].“推”指狩猎采集人群受到环境变化或生计压力被迫转向定居生活,一方面强调气候变化因素的影响,如干旱化导致的资源萎缩促使人们向环境较好的地区聚集并定居[27-28],另一方面强调人口增长导致的资源压力增加[2,29 -30],促使人群向定居转变.还有一种解释认为,区域性的人口增长导致各群体内部联系更加紧密,在当地资源枯竭时更难迁移从而驱动定居.随后定居在群体间扩散,进一步通过提高生育率促进人口增长,不断正反馈强化“定居—人口增长—定居”这一过程,使定居程度不断加深[13,31].“海洋假说”(Maritime Hypothesis)认为,一些以海洋资源为经济基础的社会可能出现类似农业的经济集约化,并趋于定居生活,并且也像以农业为基础的社会一样具有社会复杂性[1];另外,地理环境差异造成的资源不均匀分布会促使狩猎采集人群降低移动性,趋于定居,生态交错带等资源关键区则发挥了关键作用[7,32 -33].从经济和政治等社会因素角度来看,人口增长、食物储藏技术的发展、领地意识的产生、个人财产权的出现使社会连结更加紧密,人们主动或被动的加入定居的生活方式中,以分散个体所承担的风险、增加合作、提高资源获取效率、促进技术发展和人口增长以应对环境变化[9,13,20,22,32,34⇓⇓⇓⇓⇓⇓ -41].随后定居的生活方式在群体间扩散,随着不断正反馈强化,使定居程度不断加深[13,31]. ...
... ,22,32,34⇓⇓⇓⇓⇓⇓ -41].随后定居的生活方式在群体间扩散,随着不断正反馈强化,使定居程度不断加深[13,31]. ...
1
1985
... 而定居的产生则被归结为气候变化、人口增长、资源压力、技术革新等自然环境和社会经济因素共同作用的结果[21-22].“推拉假说”("Push" and "Pull" Hypotheses)强调丰富的自然资源或由干旱化等因素造成的环境压力以“拉”或“推”的方式促使人们向环境较好的地区聚集并定居[23].根据“拉”理论的假设,丰富的资源吸引人们减少流动性进而产生定居,比如非洲北部地区新石器时代早期,常年或季节性河流提供的丰富渔猎资源对狩猎采集者的吸引、纳图夫文化时期先民对瞪羚(Gazella gazella)等非长距迁徙动物和季节性植物资源利用,促使非农业狩猎采集社会的移动性降低、定居村落产生[24⇓-26].“推”指狩猎采集人群受到环境变化或生计压力被迫转向定居生活,一方面强调气候变化因素的影响,如干旱化导致的资源萎缩促使人们向环境较好的地区聚集并定居[27-28],另一方面强调人口增长导致的资源压力增加[2,29 -30],促使人群向定居转变.还有一种解释认为,区域性的人口增长导致各群体内部联系更加紧密,在当地资源枯竭时更难迁移从而驱动定居.随后定居在群体间扩散,进一步通过提高生育率促进人口增长,不断正反馈强化“定居—人口增长—定居”这一过程,使定居程度不断加深[13,31].“海洋假说”(Maritime Hypothesis)认为,一些以海洋资源为经济基础的社会可能出现类似农业的经济集约化,并趋于定居生活,并且也像以农业为基础的社会一样具有社会复杂性[1];另外,地理环境差异造成的资源不均匀分布会促使狩猎采集人群降低移动性,趋于定居,生态交错带等资源关键区则发挥了关键作用[7,32 -33].从经济和政治等社会因素角度来看,人口增长、食物储藏技术的发展、领地意识的产生、个人财产权的出现使社会连结更加紧密,人们主动或被动的加入定居的生活方式中,以分散个体所承担的风险、增加合作、提高资源获取效率、促进技术发展和人口增长以应对环境变化[9,13,20,22,32,34⇓⇓⇓⇓⇓⇓ -41].随后定居的生活方式在群体间扩散,随着不断正反馈强化,使定居程度不断加深[13,31]. ...
Semi-permanent foragers in semi-arid environments of North Africa
1
2006
... 而定居的产生则被归结为气候变化、人口增长、资源压力、技术革新等自然环境和社会经济因素共同作用的结果[21-22].“推拉假说”("Push" and "Pull" Hypotheses)强调丰富的自然资源或由干旱化等因素造成的环境压力以“拉”或“推”的方式促使人们向环境较好的地区聚集并定居[23].根据“拉”理论的假设,丰富的资源吸引人们减少流动性进而产生定居,比如非洲北部地区新石器时代早期,常年或季节性河流提供的丰富渔猎资源对狩猎采集者的吸引、纳图夫文化时期先民对瞪羚(Gazella gazella)等非长距迁徙动物和季节性植物资源利用,促使非农业狩猎采集社会的移动性降低、定居村落产生[24⇓-26].“推”指狩猎采集人群受到环境变化或生计压力被迫转向定居生活,一方面强调气候变化因素的影响,如干旱化导致的资源萎缩促使人们向环境较好的地区聚集并定居[27-28],另一方面强调人口增长导致的资源压力增加[2,29 -30],促使人群向定居转变.还有一种解释认为,区域性的人口增长导致各群体内部联系更加紧密,在当地资源枯竭时更难迁移从而驱动定居.随后定居在群体间扩散,进一步通过提高生育率促进人口增长,不断正反馈强化“定居—人口增长—定居”这一过程,使定居程度不断加深[13,31].“海洋假说”(Maritime Hypothesis)认为,一些以海洋资源为经济基础的社会可能出现类似农业的经济集约化,并趋于定居生活,并且也像以农业为基础的社会一样具有社会复杂性[1];另外,地理环境差异造成的资源不均匀分布会促使狩猎采集人群降低移动性,趋于定居,生态交错带等资源关键区则发挥了关键作用[7,32 -33].从经济和政治等社会因素角度来看,人口增长、食物储藏技术的发展、领地意识的产生、个人财产权的出现使社会连结更加紧密,人们主动或被动的加入定居的生活方式中,以分散个体所承担的风险、增加合作、提高资源获取效率、促进技术发展和人口增长以应对环境变化[9,13,20,22,32,34⇓⇓⇓⇓⇓⇓ -41].随后定居的生活方式在群体间扩散,随着不断正反馈强化,使定居程度不断加深[13,31]. ...
Reproductive strategies in female mountain and dorcas gazelles (Gazella gazella gazella and Gazella dorcas)
1
1983
... 而定居的产生则被归结为气候变化、人口增长、资源压力、技术革新等自然环境和社会经济因素共同作用的结果[21-22].“推拉假说”("Push" and "Pull" Hypotheses)强调丰富的自然资源或由干旱化等因素造成的环境压力以“拉”或“推”的方式促使人们向环境较好的地区聚集并定居[23].根据“拉”理论的假设,丰富的资源吸引人们减少流动性进而产生定居,比如非洲北部地区新石器时代早期,常年或季节性河流提供的丰富渔猎资源对狩猎采集者的吸引、纳图夫文化时期先民对瞪羚(Gazella gazella)等非长距迁徙动物和季节性植物资源利用,促使非农业狩猎采集社会的移动性降低、定居村落产生[24⇓-26].“推”指狩猎采集人群受到环境变化或生计压力被迫转向定居生活,一方面强调气候变化因素的影响,如干旱化导致的资源萎缩促使人们向环境较好的地区聚集并定居[27-28],另一方面强调人口增长导致的资源压力增加[2,29 -30],促使人群向定居转变.还有一种解释认为,区域性的人口增长导致各群体内部联系更加紧密,在当地资源枯竭时更难迁移从而驱动定居.随后定居在群体间扩散,进一步通过提高生育率促进人口增长,不断正反馈强化“定居—人口增长—定居”这一过程,使定居程度不断加深[13,31].“海洋假说”(Maritime Hypothesis)认为,一些以海洋资源为经济基础的社会可能出现类似农业的经济集约化,并趋于定居生活,并且也像以农业为基础的社会一样具有社会复杂性[1];另外,地理环境差异造成的资源不均匀分布会促使狩猎采集人群降低移动性,趋于定居,生态交错带等资源关键区则发挥了关键作用[7,32 -33].从经济和政治等社会因素角度来看,人口增长、食物储藏技术的发展、领地意识的产生、个人财产权的出现使社会连结更加紧密,人们主动或被动的加入定居的生活方式中,以分散个体所承担的风险、增加合作、提高资源获取效率、促进技术发展和人口增长以应对环境变化[9,13,20,22,32,34⇓⇓⇓⇓⇓⇓ -41].随后定居的生活方式在群体间扩散,随着不断正反馈强化,使定居程度不断加深[13,31]. ...
1
2013
... 而定居的产生则被归结为气候变化、人口增长、资源压力、技术革新等自然环境和社会经济因素共同作用的结果[21-22].“推拉假说”("Push" and "Pull" Hypotheses)强调丰富的自然资源或由干旱化等因素造成的环境压力以“拉”或“推”的方式促使人们向环境较好的地区聚集并定居[23].根据“拉”理论的假设,丰富的资源吸引人们减少流动性进而产生定居,比如非洲北部地区新石器时代早期,常年或季节性河流提供的丰富渔猎资源对狩猎采集者的吸引、纳图夫文化时期先民对瞪羚(Gazella gazella)等非长距迁徙动物和季节性植物资源利用,促使非农业狩猎采集社会的移动性降低、定居村落产生[24⇓-26].“推”指狩猎采集人群受到环境变化或生计压力被迫转向定居生活,一方面强调气候变化因素的影响,如干旱化导致的资源萎缩促使人们向环境较好的地区聚集并定居[27-28],另一方面强调人口增长导致的资源压力增加[2,29 -30],促使人群向定居转变.还有一种解释认为,区域性的人口增长导致各群体内部联系更加紧密,在当地资源枯竭时更难迁移从而驱动定居.随后定居在群体间扩散,进一步通过提高生育率促进人口增长,不断正反馈强化“定居—人口增长—定居”这一过程,使定居程度不断加深[13,31].“海洋假说”(Maritime Hypothesis)认为,一些以海洋资源为经济基础的社会可能出现类似农业的经济集约化,并趋于定居生活,并且也像以农业为基础的社会一样具有社会复杂性[1];另外,地理环境差异造成的资源不均匀分布会促使狩猎采集人群降低移动性,趋于定居,生态交错带等资源关键区则发挥了关键作用[7,32 -33].从经济和政治等社会因素角度来看,人口增长、食物储藏技术的发展、领地意识的产生、个人财产权的出现使社会连结更加紧密,人们主动或被动的加入定居的生活方式中,以分散个体所承担的风险、增加合作、提高资源获取效率、促进技术发展和人口增长以应对环境变化[9,13,20,22,32,34⇓⇓⇓⇓⇓⇓ -41].随后定居的生活方式在群体间扩散,随着不断正反馈强化,使定居程度不断加深[13,31]. ...
The origins of sedentism and farming communities in the Levant
1
1989
... 而定居的产生则被归结为气候变化、人口增长、资源压力、技术革新等自然环境和社会经济因素共同作用的结果[21-22].“推拉假说”("Push" and "Pull" Hypotheses)强调丰富的自然资源或由干旱化等因素造成的环境压力以“拉”或“推”的方式促使人们向环境较好的地区聚集并定居[23].根据“拉”理论的假设,丰富的资源吸引人们减少流动性进而产生定居,比如非洲北部地区新石器时代早期,常年或季节性河流提供的丰富渔猎资源对狩猎采集者的吸引、纳图夫文化时期先民对瞪羚(Gazella gazella)等非长距迁徙动物和季节性植物资源利用,促使非农业狩猎采集社会的移动性降低、定居村落产生[24⇓-26].“推”指狩猎采集人群受到环境变化或生计压力被迫转向定居生活,一方面强调气候变化因素的影响,如干旱化导致的资源萎缩促使人们向环境较好的地区聚集并定居[27-28],另一方面强调人口增长导致的资源压力增加[2,29 -30],促使人群向定居转变.还有一种解释认为,区域性的人口增长导致各群体内部联系更加紧密,在当地资源枯竭时更难迁移从而驱动定居.随后定居在群体间扩散,进一步通过提高生育率促进人口增长,不断正反馈强化“定居—人口增长—定居”这一过程,使定居程度不断加深[13,31].“海洋假说”(Maritime Hypothesis)认为,一些以海洋资源为经济基础的社会可能出现类似农业的经济集约化,并趋于定居生活,并且也像以农业为基础的社会一样具有社会复杂性[1];另外,地理环境差异造成的资源不均匀分布会促使狩猎采集人群降低移动性,趋于定居,生态交错带等资源关键区则发挥了关键作用[7,32 -33].从经济和政治等社会因素角度来看,人口增长、食物储藏技术的发展、领地意识的产生、个人财产权的出现使社会连结更加紧密,人们主动或被动的加入定居的生活方式中,以分散个体所承担的风险、增加合作、提高资源获取效率、促进技术发展和人口增长以应对环境变化[9,13,20,22,32,34⇓⇓⇓⇓⇓⇓ -41].随后定居的生活方式在群体间扩散,随着不断正反馈强化,使定居程度不断加深[13,31]. ...
1
2008
... 而定居的产生则被归结为气候变化、人口增长、资源压力、技术革新等自然环境和社会经济因素共同作用的结果[21-22].“推拉假说”("Push" and "Pull" Hypotheses)强调丰富的自然资源或由干旱化等因素造成的环境压力以“拉”或“推”的方式促使人们向环境较好的地区聚集并定居[23].根据“拉”理论的假设,丰富的资源吸引人们减少流动性进而产生定居,比如非洲北部地区新石器时代早期,常年或季节性河流提供的丰富渔猎资源对狩猎采集者的吸引、纳图夫文化时期先民对瞪羚(Gazella gazella)等非长距迁徙动物和季节性植物资源利用,促使非农业狩猎采集社会的移动性降低、定居村落产生[24⇓-26].“推”指狩猎采集人群受到环境变化或生计压力被迫转向定居生活,一方面强调气候变化因素的影响,如干旱化导致的资源萎缩促使人们向环境较好的地区聚集并定居[27-28],另一方面强调人口增长导致的资源压力增加[2,29 -30],促使人群向定居转变.还有一种解释认为,区域性的人口增长导致各群体内部联系更加紧密,在当地资源枯竭时更难迁移从而驱动定居.随后定居在群体间扩散,进一步通过提高生育率促进人口增长,不断正反馈强化“定居—人口增长—定居”这一过程,使定居程度不断加深[13,31].“海洋假说”(Maritime Hypothesis)认为,一些以海洋资源为经济基础的社会可能出现类似农业的经济集约化,并趋于定居生活,并且也像以农业为基础的社会一样具有社会复杂性[1];另外,地理环境差异造成的资源不均匀分布会促使狩猎采集人群降低移动性,趋于定居,生态交错带等资源关键区则发挥了关键作用[7,32 -33].从经济和政治等社会因素角度来看,人口增长、食物储藏技术的发展、领地意识的产生、个人财产权的出现使社会连结更加紧密,人们主动或被动的加入定居的生活方式中,以分散个体所承担的风险、增加合作、提高资源获取效率、促进技术发展和人口增长以应对环境变化[9,13,20,22,32,34⇓⇓⇓⇓⇓⇓ -41].随后定居的生活方式在群体间扩散,随着不断正反馈强化,使定居程度不断加深[13,31]. ...
1
1985
... 而定居的产生则被归结为气候变化、人口增长、资源压力、技术革新等自然环境和社会经济因素共同作用的结果[21-22].“推拉假说”("Push" and "Pull" Hypotheses)强调丰富的自然资源或由干旱化等因素造成的环境压力以“拉”或“推”的方式促使人们向环境较好的地区聚集并定居[23].根据“拉”理论的假设,丰富的资源吸引人们减少流动性进而产生定居,比如非洲北部地区新石器时代早期,常年或季节性河流提供的丰富渔猎资源对狩猎采集者的吸引、纳图夫文化时期先民对瞪羚(Gazella gazella)等非长距迁徙动物和季节性植物资源利用,促使非农业狩猎采集社会的移动性降低、定居村落产生[24⇓-26].“推”指狩猎采集人群受到环境变化或生计压力被迫转向定居生活,一方面强调气候变化因素的影响,如干旱化导致的资源萎缩促使人们向环境较好的地区聚集并定居[27-28],另一方面强调人口增长导致的资源压力增加[2,29 -30],促使人群向定居转变.还有一种解释认为,区域性的人口增长导致各群体内部联系更加紧密,在当地资源枯竭时更难迁移从而驱动定居.随后定居在群体间扩散,进一步通过提高生育率促进人口增长,不断正反馈强化“定居—人口增长—定居”这一过程,使定居程度不断加深[13,31].“海洋假说”(Maritime Hypothesis)认为,一些以海洋资源为经济基础的社会可能出现类似农业的经济集约化,并趋于定居生活,并且也像以农业为基础的社会一样具有社会复杂性[1];另外,地理环境差异造成的资源不均匀分布会促使狩猎采集人群降低移动性,趋于定居,生态交错带等资源关键区则发挥了关键作用[7,32 -33].从经济和政治等社会因素角度来看,人口增长、食物储藏技术的发展、领地意识的产生、个人财产权的出现使社会连结更加紧密,人们主动或被动的加入定居的生活方式中,以分散个体所承担的风险、增加合作、提高资源获取效率、促进技术发展和人口增长以应对环境变化[9,13,20,22,32,34⇓⇓⇓⇓⇓⇓ -41].随后定居的生活方式在群体间扩散,随着不断正反馈强化,使定居程度不断加深[13,31]. ...
Sedentism: A temporal shift or a transitional change in hunter-gatherer mobility patterns? Between Bands and States
1
... 而定居的产生则被归结为气候变化、人口增长、资源压力、技术革新等自然环境和社会经济因素共同作用的结果[21-22].“推拉假说”("Push" and "Pull" Hypotheses)强调丰富的自然资源或由干旱化等因素造成的环境压力以“拉”或“推”的方式促使人们向环境较好的地区聚集并定居[23].根据“拉”理论的假设,丰富的资源吸引人们减少流动性进而产生定居,比如非洲北部地区新石器时代早期,常年或季节性河流提供的丰富渔猎资源对狩猎采集者的吸引、纳图夫文化时期先民对瞪羚(Gazella gazella)等非长距迁徙动物和季节性植物资源利用,促使非农业狩猎采集社会的移动性降低、定居村落产生[24⇓-26].“推”指狩猎采集人群受到环境变化或生计压力被迫转向定居生活,一方面强调气候变化因素的影响,如干旱化导致的资源萎缩促使人们向环境较好的地区聚集并定居[27-28],另一方面强调人口增长导致的资源压力增加[2,29 -30],促使人群向定居转变.还有一种解释认为,区域性的人口增长导致各群体内部联系更加紧密,在当地资源枯竭时更难迁移从而驱动定居.随后定居在群体间扩散,进一步通过提高生育率促进人口增长,不断正反馈强化“定居—人口增长—定居”这一过程,使定居程度不断加深[13,31].“海洋假说”(Maritime Hypothesis)认为,一些以海洋资源为经济基础的社会可能出现类似农业的经济集约化,并趋于定居生活,并且也像以农业为基础的社会一样具有社会复杂性[1];另外,地理环境差异造成的资源不均匀分布会促使狩猎采集人群降低移动性,趋于定居,生态交错带等资源关键区则发挥了关键作用[7,32 -33].从经济和政治等社会因素角度来看,人口增长、食物储藏技术的发展、领地意识的产生、个人财产权的出现使社会连结更加紧密,人们主动或被动的加入定居的生活方式中,以分散个体所承担的风险、增加合作、提高资源获取效率、促进技术发展和人口增长以应对环境变化[9,13,20,22,32,34⇓⇓⇓⇓⇓⇓ -41].随后定居的生活方式在群体间扩散,随着不断正反馈强化,使定居程度不断加深[13,31]. ...
From the peaceful to the warlike: Ethnographic and archaeological insights into hunter-gatherer warfare and homicide//Fry D P. War, Peace, and Human Nature: The Convergence of Evolutionary and Cultural Views
2
2013
... 而定居的产生则被归结为气候变化、人口增长、资源压力、技术革新等自然环境和社会经济因素共同作用的结果[21-22].“推拉假说”("Push" and "Pull" Hypotheses)强调丰富的自然资源或由干旱化等因素造成的环境压力以“拉”或“推”的方式促使人们向环境较好的地区聚集并定居[23].根据“拉”理论的假设,丰富的资源吸引人们减少流动性进而产生定居,比如非洲北部地区新石器时代早期,常年或季节性河流提供的丰富渔猎资源对狩猎采集者的吸引、纳图夫文化时期先民对瞪羚(Gazella gazella)等非长距迁徙动物和季节性植物资源利用,促使非农业狩猎采集社会的移动性降低、定居村落产生[24⇓-26].“推”指狩猎采集人群受到环境变化或生计压力被迫转向定居生活,一方面强调气候变化因素的影响,如干旱化导致的资源萎缩促使人们向环境较好的地区聚集并定居[27-28],另一方面强调人口增长导致的资源压力增加[2,29 -30],促使人群向定居转变.还有一种解释认为,区域性的人口增长导致各群体内部联系更加紧密,在当地资源枯竭时更难迁移从而驱动定居.随后定居在群体间扩散,进一步通过提高生育率促进人口增长,不断正反馈强化“定居—人口增长—定居”这一过程,使定居程度不断加深[13,31].“海洋假说”(Maritime Hypothesis)认为,一些以海洋资源为经济基础的社会可能出现类似农业的经济集约化,并趋于定居生活,并且也像以农业为基础的社会一样具有社会复杂性[1];另外,地理环境差异造成的资源不均匀分布会促使狩猎采集人群降低移动性,趋于定居,生态交错带等资源关键区则发挥了关键作用[7,32 -33].从经济和政治等社会因素角度来看,人口增长、食物储藏技术的发展、领地意识的产生、个人财产权的出现使社会连结更加紧密,人们主动或被动的加入定居的生活方式中,以分散个体所承担的风险、增加合作、提高资源获取效率、促进技术发展和人口增长以应对环境变化[9,13,20,22,32,34⇓⇓⇓⇓⇓⇓ -41].随后定居的生活方式在群体间扩散,随着不断正反馈强化,使定居程度不断加深[13,31]. ...
... ,31]. ...
Willow smoke and dogs' tails: Hunter-gatherer settlement systems and archaeological site formation
2
1980
... 而定居的产生则被归结为气候变化、人口增长、资源压力、技术革新等自然环境和社会经济因素共同作用的结果[21-22].“推拉假说”("Push" and "Pull" Hypotheses)强调丰富的自然资源或由干旱化等因素造成的环境压力以“拉”或“推”的方式促使人们向环境较好的地区聚集并定居[23].根据“拉”理论的假设,丰富的资源吸引人们减少流动性进而产生定居,比如非洲北部地区新石器时代早期,常年或季节性河流提供的丰富渔猎资源对狩猎采集者的吸引、纳图夫文化时期先民对瞪羚(Gazella gazella)等非长距迁徙动物和季节性植物资源利用,促使非农业狩猎采集社会的移动性降低、定居村落产生[24⇓-26].“推”指狩猎采集人群受到环境变化或生计压力被迫转向定居生活,一方面强调气候变化因素的影响,如干旱化导致的资源萎缩促使人们向环境较好的地区聚集并定居[27-28],另一方面强调人口增长导致的资源压力增加[2,29 -30],促使人群向定居转变.还有一种解释认为,区域性的人口增长导致各群体内部联系更加紧密,在当地资源枯竭时更难迁移从而驱动定居.随后定居在群体间扩散,进一步通过提高生育率促进人口增长,不断正反馈强化“定居—人口增长—定居”这一过程,使定居程度不断加深[13,31].“海洋假说”(Maritime Hypothesis)认为,一些以海洋资源为经济基础的社会可能出现类似农业的经济集约化,并趋于定居生活,并且也像以农业为基础的社会一样具有社会复杂性[1];另外,地理环境差异造成的资源不均匀分布会促使狩猎采集人群降低移动性,趋于定居,生态交错带等资源关键区则发挥了关键作用[7,32 -33].从经济和政治等社会因素角度来看,人口增长、食物储藏技术的发展、领地意识的产生、个人财产权的出现使社会连结更加紧密,人们主动或被动的加入定居的生活方式中,以分散个体所承担的风险、增加合作、提高资源获取效率、促进技术发展和人口增长以应对环境变化[9,13,20,22,32,34⇓⇓⇓⇓⇓⇓ -41].随后定居的生活方式在群体间扩散,随着不断正反馈强化,使定居程度不断加深[13,31]. ...
... ,32,34⇓⇓⇓⇓⇓⇓ -41].随后定居的生活方式在群体间扩散,随着不断正反馈强化,使定居程度不断加深[13,31]. ...
1
1968
... 而定居的产生则被归结为气候变化、人口增长、资源压力、技术革新等自然环境和社会经济因素共同作用的结果[21-22].“推拉假说”("Push" and "Pull" Hypotheses)强调丰富的自然资源或由干旱化等因素造成的环境压力以“拉”或“推”的方式促使人们向环境较好的地区聚集并定居[23].根据“拉”理论的假设,丰富的资源吸引人们减少流动性进而产生定居,比如非洲北部地区新石器时代早期,常年或季节性河流提供的丰富渔猎资源对狩猎采集者的吸引、纳图夫文化时期先民对瞪羚(Gazella gazella)等非长距迁徙动物和季节性植物资源利用,促使非农业狩猎采集社会的移动性降低、定居村落产生[24⇓-26].“推”指狩猎采集人群受到环境变化或生计压力被迫转向定居生活,一方面强调气候变化因素的影响,如干旱化导致的资源萎缩促使人们向环境较好的地区聚集并定居[27-28],另一方面强调人口增长导致的资源压力增加[2,29 -30],促使人群向定居转变.还有一种解释认为,区域性的人口增长导致各群体内部联系更加紧密,在当地资源枯竭时更难迁移从而驱动定居.随后定居在群体间扩散,进一步通过提高生育率促进人口增长,不断正反馈强化“定居—人口增长—定居”这一过程,使定居程度不断加深[13,31].“海洋假说”(Maritime Hypothesis)认为,一些以海洋资源为经济基础的社会可能出现类似农业的经济集约化,并趋于定居生活,并且也像以农业为基础的社会一样具有社会复杂性[1];另外,地理环境差异造成的资源不均匀分布会促使狩猎采集人群降低移动性,趋于定居,生态交错带等资源关键区则发挥了关键作用[7,32 -33].从经济和政治等社会因素角度来看,人口增长、食物储藏技术的发展、领地意识的产生、个人财产权的出现使社会连结更加紧密,人们主动或被动的加入定居的生活方式中,以分散个体所承担的风险、增加合作、提高资源获取效率、促进技术发展和人口增长以应对环境变化[9,13,20,22,32,34⇓⇓⇓⇓⇓⇓ -41].随后定居的生活方式在群体间扩散,随着不断正反馈强化,使定居程度不断加深[13,31]. ...
Some thoughts on the Middle to Upper Paleolithic transition
1
1982
... 而定居的产生则被归结为气候变化、人口增长、资源压力、技术革新等自然环境和社会经济因素共同作用的结果[21-22].“推拉假说”("Push" and "Pull" Hypotheses)强调丰富的自然资源或由干旱化等因素造成的环境压力以“拉”或“推”的方式促使人们向环境较好的地区聚集并定居[23].根据“拉”理论的假设,丰富的资源吸引人们减少流动性进而产生定居,比如非洲北部地区新石器时代早期,常年或季节性河流提供的丰富渔猎资源对狩猎采集者的吸引、纳图夫文化时期先民对瞪羚(Gazella gazella)等非长距迁徙动物和季节性植物资源利用,促使非农业狩猎采集社会的移动性降低、定居村落产生[24⇓-26].“推”指狩猎采集人群受到环境变化或生计压力被迫转向定居生活,一方面强调气候变化因素的影响,如干旱化导致的资源萎缩促使人们向环境较好的地区聚集并定居[27-28],另一方面强调人口增长导致的资源压力增加[2,29 -30],促使人群向定居转变.还有一种解释认为,区域性的人口增长导致各群体内部联系更加紧密,在当地资源枯竭时更难迁移从而驱动定居.随后定居在群体间扩散,进一步通过提高生育率促进人口增长,不断正反馈强化“定居—人口增长—定居”这一过程,使定居程度不断加深[13,31].“海洋假说”(Maritime Hypothesis)认为,一些以海洋资源为经济基础的社会可能出现类似农业的经济集约化,并趋于定居生活,并且也像以农业为基础的社会一样具有社会复杂性[1];另外,地理环境差异造成的资源不均匀分布会促使狩猎采集人群降低移动性,趋于定居,生态交错带等资源关键区则发挥了关键作用[7,32 -33].从经济和政治等社会因素角度来看,人口增长、食物储藏技术的发展、领地意识的产生、个人财产权的出现使社会连结更加紧密,人们主动或被动的加入定居的生活方式中,以分散个体所承担的风险、增加合作、提高资源获取效率、促进技术发展和人口增长以应对环境变化[9,13,20,22,32,34⇓⇓⇓⇓⇓⇓ -41].随后定居的生活方式在群体间扩散,随着不断正反馈强化,使定居程度不断加深[13,31]. ...
1
2002
... 而定居的产生则被归结为气候变化、人口增长、资源压力、技术革新等自然环境和社会经济因素共同作用的结果[21-22].“推拉假说”("Push" and "Pull" Hypotheses)强调丰富的自然资源或由干旱化等因素造成的环境压力以“拉”或“推”的方式促使人们向环境较好的地区聚集并定居[23].根据“拉”理论的假设,丰富的资源吸引人们减少流动性进而产生定居,比如非洲北部地区新石器时代早期,常年或季节性河流提供的丰富渔猎资源对狩猎采集者的吸引、纳图夫文化时期先民对瞪羚(Gazella gazella)等非长距迁徙动物和季节性植物资源利用,促使非农业狩猎采集社会的移动性降低、定居村落产生[24⇓-26].“推”指狩猎采集人群受到环境变化或生计压力被迫转向定居生活,一方面强调气候变化因素的影响,如干旱化导致的资源萎缩促使人们向环境较好的地区聚集并定居[27-28],另一方面强调人口增长导致的资源压力增加[2,29 -30],促使人群向定居转变.还有一种解释认为,区域性的人口增长导致各群体内部联系更加紧密,在当地资源枯竭时更难迁移从而驱动定居.随后定居在群体间扩散,进一步通过提高生育率促进人口增长,不断正反馈强化“定居—人口增长—定居”这一过程,使定居程度不断加深[13,31].“海洋假说”(Maritime Hypothesis)认为,一些以海洋资源为经济基础的社会可能出现类似农业的经济集约化,并趋于定居生活,并且也像以农业为基础的社会一样具有社会复杂性[1];另外,地理环境差异造成的资源不均匀分布会促使狩猎采集人群降低移动性,趋于定居,生态交错带等资源关键区则发挥了关键作用[7,32 -33].从经济和政治等社会因素角度来看,人口增长、食物储藏技术的发展、领地意识的产生、个人财产权的出现使社会连结更加紧密,人们主动或被动的加入定居的生活方式中,以分散个体所承担的风险、增加合作、提高资源获取效率、促进技术发展和人口增长以应对环境变化[9,13,20,22,32,34⇓⇓⇓⇓⇓⇓ -41].随后定居的生活方式在群体间扩散,随着不断正反馈强化,使定居程度不断加深[13,31]. ...
Social Organisation and Settlement:Contributions from Anthropology
1
... 而定居的产生则被归结为气候变化、人口增长、资源压力、技术革新等自然环境和社会经济因素共同作用的结果[21-22].“推拉假说”("Push" and "Pull" Hypotheses)强调丰富的自然资源或由干旱化等因素造成的环境压力以“拉”或“推”的方式促使人们向环境较好的地区聚集并定居[23].根据“拉”理论的假设,丰富的资源吸引人们减少流动性进而产生定居,比如非洲北部地区新石器时代早期,常年或季节性河流提供的丰富渔猎资源对狩猎采集者的吸引、纳图夫文化时期先民对瞪羚(Gazella gazella)等非长距迁徙动物和季节性植物资源利用,促使非农业狩猎采集社会的移动性降低、定居村落产生[24⇓-26].“推”指狩猎采集人群受到环境变化或生计压力被迫转向定居生活,一方面强调气候变化因素的影响,如干旱化导致的资源萎缩促使人们向环境较好的地区聚集并定居[27-28],另一方面强调人口增长导致的资源压力增加[2,29 -30],促使人群向定居转变.还有一种解释认为,区域性的人口增长导致各群体内部联系更加紧密,在当地资源枯竭时更难迁移从而驱动定居.随后定居在群体间扩散,进一步通过提高生育率促进人口增长,不断正反馈强化“定居—人口增长—定居”这一过程,使定居程度不断加深[13,31].“海洋假说”(Maritime Hypothesis)认为,一些以海洋资源为经济基础的社会可能出现类似农业的经济集约化,并趋于定居生活,并且也像以农业为基础的社会一样具有社会复杂性[1];另外,地理环境差异造成的资源不均匀分布会促使狩猎采集人群降低移动性,趋于定居,生态交错带等资源关键区则发挥了关键作用[7,32 -33].从经济和政治等社会因素角度来看,人口增长、食物储藏技术的发展、领地意识的产生、个人财产权的出现使社会连结更加紧密,人们主动或被动的加入定居的生活方式中,以分散个体所承担的风险、增加合作、提高资源获取效率、促进技术发展和人口增长以应对环境变化[9,13,20,22,32,34⇓⇓⇓⇓⇓⇓ -41].随后定居的生活方式在群体间扩散,随着不断正反馈强化,使定居程度不断加深[13,31]. ...
1
1982
... 而定居的产生则被归结为气候变化、人口增长、资源压力、技术革新等自然环境和社会经济因素共同作用的结果[21-22].“推拉假说”("Push" and "Pull" Hypotheses)强调丰富的自然资源或由干旱化等因素造成的环境压力以“拉”或“推”的方式促使人们向环境较好的地区聚集并定居[23].根据“拉”理论的假设,丰富的资源吸引人们减少流动性进而产生定居,比如非洲北部地区新石器时代早期,常年或季节性河流提供的丰富渔猎资源对狩猎采集者的吸引、纳图夫文化时期先民对瞪羚(Gazella gazella)等非长距迁徙动物和季节性植物资源利用,促使非农业狩猎采集社会的移动性降低、定居村落产生[24⇓-26].“推”指狩猎采集人群受到环境变化或生计压力被迫转向定居生活,一方面强调气候变化因素的影响,如干旱化导致的资源萎缩促使人们向环境较好的地区聚集并定居[27-28],另一方面强调人口增长导致的资源压力增加[2,29 -30],促使人群向定居转变.还有一种解释认为,区域性的人口增长导致各群体内部联系更加紧密,在当地资源枯竭时更难迁移从而驱动定居.随后定居在群体间扩散,进一步通过提高生育率促进人口增长,不断正反馈强化“定居—人口增长—定居”这一过程,使定居程度不断加深[13,31].“海洋假说”(Maritime Hypothesis)认为,一些以海洋资源为经济基础的社会可能出现类似农业的经济集约化,并趋于定居生活,并且也像以农业为基础的社会一样具有社会复杂性[1];另外,地理环境差异造成的资源不均匀分布会促使狩猎采集人群降低移动性,趋于定居,生态交错带等资源关键区则发挥了关键作用[7,32 -33].从经济和政治等社会因素角度来看,人口增长、食物储藏技术的发展、领地意识的产生、个人财产权的出现使社会连结更加紧密,人们主动或被动的加入定居的生活方式中,以分散个体所承担的风险、增加合作、提高资源获取效率、促进技术发展和人口增长以应对环境变化[9,13,20,22,32,34⇓⇓⇓⇓⇓⇓ -41].随后定居的生活方式在群体间扩散,随着不断正反馈强化,使定居程度不断加深[13,31]. ...
An equilibrium conflict model of land tenure in hunter-gatherer societies
1
2003
... 而定居的产生则被归结为气候变化、人口增长、资源压力、技术革新等自然环境和社会经济因素共同作用的结果[21-22].“推拉假说”("Push" and "Pull" Hypotheses)强调丰富的自然资源或由干旱化等因素造成的环境压力以“拉”或“推”的方式促使人们向环境较好的地区聚集并定居[23].根据“拉”理论的假设,丰富的资源吸引人们减少流动性进而产生定居,比如非洲北部地区新石器时代早期,常年或季节性河流提供的丰富渔猎资源对狩猎采集者的吸引、纳图夫文化时期先民对瞪羚(Gazella gazella)等非长距迁徙动物和季节性植物资源利用,促使非农业狩猎采集社会的移动性降低、定居村落产生[24⇓-26].“推”指狩猎采集人群受到环境变化或生计压力被迫转向定居生活,一方面强调气候变化因素的影响,如干旱化导致的资源萎缩促使人们向环境较好的地区聚集并定居[27-28],另一方面强调人口增长导致的资源压力增加[2,29 -30],促使人群向定居转变.还有一种解释认为,区域性的人口增长导致各群体内部联系更加紧密,在当地资源枯竭时更难迁移从而驱动定居.随后定居在群体间扩散,进一步通过提高生育率促进人口增长,不断正反馈强化“定居—人口增长—定居”这一过程,使定居程度不断加深[13,31].“海洋假说”(Maritime Hypothesis)认为,一些以海洋资源为经济基础的社会可能出现类似农业的经济集约化,并趋于定居生活,并且也像以农业为基础的社会一样具有社会复杂性[1];另外,地理环境差异造成的资源不均匀分布会促使狩猎采集人群降低移动性,趋于定居,生态交错带等资源关键区则发挥了关键作用[7,32 -33].从经济和政治等社会因素角度来看,人口增长、食物储藏技术的发展、领地意识的产生、个人财产权的出现使社会连结更加紧密,人们主动或被动的加入定居的生活方式中,以分散个体所承担的风险、增加合作、提高资源获取效率、促进技术发展和人口增长以应对环境变化[9,13,20,22,32,34⇓⇓⇓⇓⇓⇓ -41].随后定居的生活方式在群体间扩散,随着不断正反馈强化,使定居程度不断加深[13,31]. ...
Origins of food production in southwestern Asia: A survey of ideas
1
1971
... 而定居的产生则被归结为气候变化、人口增长、资源压力、技术革新等自然环境和社会经济因素共同作用的结果[21-22].“推拉假说”("Push" and "Pull" Hypotheses)强调丰富的自然资源或由干旱化等因素造成的环境压力以“拉”或“推”的方式促使人们向环境较好的地区聚集并定居[23].根据“拉”理论的假设,丰富的资源吸引人们减少流动性进而产生定居,比如非洲北部地区新石器时代早期,常年或季节性河流提供的丰富渔猎资源对狩猎采集者的吸引、纳图夫文化时期先民对瞪羚(Gazella gazella)等非长距迁徙动物和季节性植物资源利用,促使非农业狩猎采集社会的移动性降低、定居村落产生[24⇓-26].“推”指狩猎采集人群受到环境变化或生计压力被迫转向定居生活,一方面强调气候变化因素的影响,如干旱化导致的资源萎缩促使人们向环境较好的地区聚集并定居[27-28],另一方面强调人口增长导致的资源压力增加[2,29 -30],促使人群向定居转变.还有一种解释认为,区域性的人口增长导致各群体内部联系更加紧密,在当地资源枯竭时更难迁移从而驱动定居.随后定居在群体间扩散,进一步通过提高生育率促进人口增长,不断正反馈强化“定居—人口增长—定居”这一过程,使定居程度不断加深[13,31].“海洋假说”(Maritime Hypothesis)认为,一些以海洋资源为经济基础的社会可能出现类似农业的经济集约化,并趋于定居生活,并且也像以农业为基础的社会一样具有社会复杂性[1];另外,地理环境差异造成的资源不均匀分布会促使狩猎采集人群降低移动性,趋于定居,生态交错带等资源关键区则发挥了关键作用[7,32 -33].从经济和政治等社会因素角度来看,人口增长、食物储藏技术的发展、领地意识的产生、个人财产权的出现使社会连结更加紧密,人们主动或被动的加入定居的生活方式中,以分散个体所承担的风险、增加合作、提高资源获取效率、促进技术发展和人口增长以应对环境变化[9,13,20,22,32,34⇓⇓⇓⇓⇓⇓ -41].随后定居的生活方式在群体间扩散,随着不断正反馈强化,使定居程度不断加深[13,31]. ...
Sedentism, storage, and the intensification of small seeds: Prehistoric developments in Owens Valley
1
2003
... 而定居的产生则被归结为气候变化、人口增长、资源压力、技术革新等自然环境和社会经济因素共同作用的结果[21-22].“推拉假说”("Push" and "Pull" Hypotheses)强调丰富的自然资源或由干旱化等因素造成的环境压力以“拉”或“推”的方式促使人们向环境较好的地区聚集并定居[23].根据“拉”理论的假设,丰富的资源吸引人们减少流动性进而产生定居,比如非洲北部地区新石器时代早期,常年或季节性河流提供的丰富渔猎资源对狩猎采集者的吸引、纳图夫文化时期先民对瞪羚(Gazella gazella)等非长距迁徙动物和季节性植物资源利用,促使非农业狩猎采集社会的移动性降低、定居村落产生[24⇓-26].“推”指狩猎采集人群受到环境变化或生计压力被迫转向定居生活,一方面强调气候变化因素的影响,如干旱化导致的资源萎缩促使人们向环境较好的地区聚集并定居[27-28],另一方面强调人口增长导致的资源压力增加[2,29 -30],促使人群向定居转变.还有一种解释认为,区域性的人口增长导致各群体内部联系更加紧密,在当地资源枯竭时更难迁移从而驱动定居.随后定居在群体间扩散,进一步通过提高生育率促进人口增长,不断正反馈强化“定居—人口增长—定居”这一过程,使定居程度不断加深[13,31].“海洋假说”(Maritime Hypothesis)认为,一些以海洋资源为经济基础的社会可能出现类似农业的经济集约化,并趋于定居生活,并且也像以农业为基础的社会一样具有社会复杂性[1];另外,地理环境差异造成的资源不均匀分布会促使狩猎采集人群降低移动性,趋于定居,生态交错带等资源关键区则发挥了关键作用[7,32 -33].从经济和政治等社会因素角度来看,人口增长、食物储藏技术的发展、领地意识的产生、个人财产权的出现使社会连结更加紧密,人们主动或被动的加入定居的生活方式中,以分散个体所承担的风险、增加合作、提高资源获取效率、促进技术发展和人口增长以应对环境变化[9,13,20,22,32,34⇓⇓⇓⇓⇓⇓ -41].随后定居的生活方式在群体间扩散,随着不断正反馈强化,使定居程度不断加深[13,31]. ...
Beyond willow smoke and dogs' tails: A comment on Binford's analysis of hunter-gatherer settlement systems
1
1982
... 而定居的产生则被归结为气候变化、人口增长、资源压力、技术革新等自然环境和社会经济因素共同作用的结果[21-22].“推拉假说”("Push" and "Pull" Hypotheses)强调丰富的自然资源或由干旱化等因素造成的环境压力以“拉”或“推”的方式促使人们向环境较好的地区聚集并定居[23].根据“拉”理论的假设,丰富的资源吸引人们减少流动性进而产生定居,比如非洲北部地区新石器时代早期,常年或季节性河流提供的丰富渔猎资源对狩猎采集者的吸引、纳图夫文化时期先民对瞪羚(Gazella gazella)等非长距迁徙动物和季节性植物资源利用,促使非农业狩猎采集社会的移动性降低、定居村落产生[24⇓-26].“推”指狩猎采集人群受到环境变化或生计压力被迫转向定居生活,一方面强调气候变化因素的影响,如干旱化导致的资源萎缩促使人们向环境较好的地区聚集并定居[27-28],另一方面强调人口增长导致的资源压力增加[2,29 -30],促使人群向定居转变.还有一种解释认为,区域性的人口增长导致各群体内部联系更加紧密,在当地资源枯竭时更难迁移从而驱动定居.随后定居在群体间扩散,进一步通过提高生育率促进人口增长,不断正反馈强化“定居—人口增长—定居”这一过程,使定居程度不断加深[13,31].“海洋假说”(Maritime Hypothesis)认为,一些以海洋资源为经济基础的社会可能出现类似农业的经济集约化,并趋于定居生活,并且也像以农业为基础的社会一样具有社会复杂性[1];另外,地理环境差异造成的资源不均匀分布会促使狩猎采集人群降低移动性,趋于定居,生态交错带等资源关键区则发挥了关键作用[7,32 -33].从经济和政治等社会因素角度来看,人口增长、食物储藏技术的发展、领地意识的产生、个人财产权的出现使社会连结更加紧密,人们主动或被动的加入定居的生活方式中,以分散个体所承担的风险、增加合作、提高资源获取效率、促进技术发展和人口增长以应对环境变化[9,13,20,22,32,34⇓⇓⇓⇓⇓⇓ -41].随后定居的生活方式在群体间扩散,随着不断正反馈强化,使定居程度不断加深[13,31]. ...
2
1995
... 判断一个遗址是否出现定居有多条标准,包括投入大量人力物力建造耐用的建 筑、重型物质文化遗存如大型石磨盘、储藏设施、墓地、伴生动物如家鼠和家麻雀(Passer domesticus)的出现,动植物遗存所显示的遗址使用季节、陶器的出现和文化层厚度等[1,7,9,11,42 -43].尽管遗址往往无法符合以上全部条件,在判断遗址是否出现定居时仍需考虑多个因素. ...
... 在4.5—3.0 ka BP,史前东西方文化交流加强,驯化于西亚的黄牛、山羊(Capra hircus)和绵羊传入至青海省东北部和云南地区,青藏高原地区先民动物资源利用在空间上呈现多元化,牛和羊超过了猪,成为家畜饲养的主流(图2).家羊(绵羊/山羊)在青藏高原最早出土于金蝉口遗址(4.5—3.6 ka BP、海拔2419 m).长宁遗址(4.3—3.6 ka BP、海拔2300 m)出土的动物遗存以家畜黄牛、山羊和绵羊为主,牛、羊所占比均超过了猪,在先民生业经济中占据主要地位[42,95].青藏高原南部拉萨曲贡遗址先民的生业经济以作物种植和放牧牦牛、绵羊的农牧经济为主,以猎取白唇鹿或马鹿、麝、野猪、藏野驴和各种禽类的狩猎经济为辅[70].在高原西部阿里扎达的格布赛鲁遗址早期墓葬(3560—3000 a BP、海拔3800 m)中随葬有山羊、绵羊和黄牛等家养动物,以及盘羊和鹿等野生动物;遗址晚期墓葬(2310—2127 a BP)中出土了山羊、绵羊、黄牛、马、野驴和鱼等.在3.0 ka BP以后,绵羊、山羊、黄牛、马、狗和牦牛等家养动物在青藏高原被广泛畜养,并在人们日常生活和社会仪式中扮演重要角色[100⇓⇓-103](图2). ...
Wadi Hammeh 27: An Early Natufian Settlement at Pella in Jordan
2
2012
... 判断一个遗址是否出现定居有多条标准,包括投入大量人力物力建造耐用的建 筑、重型物质文化遗存如大型石磨盘、储藏设施、墓地、伴生动物如家鼠和家麻雀(Passer domesticus)的出现,动植物遗存所显示的遗址使用季节、陶器的出现和文化层厚度等[1,7,9,11,42 -43].尽管遗址往往无法符合以上全部条件,在判断遗址是否出现定居时仍需考虑多个因素. ...
... 季节性分析是指示遗址是否为全年居住的直接证据,也反映了人类利用自然资源 的策略和能力.温带地区动物的生长、繁殖和行为都受到季节变化的影响,因此遗址出土的动物遗存是进行遗址季节性分析的重要材料.判断方法包括但不限于:① 是否出现只在特定时间出现的物种,包括候鸟、洄游的鱼类、有迁徙习性的哺乳动物和昆虫的蛹等[107,128];② 一些骨骼的形成具有季节性特征,如鹿角生长—脱落的年度循环和雌性鸟类繁殖季节才会形成的髓质骨[104,108,129];③ 动物的出生季节相对固定,通过牙齿和骨骼判断哺乳动物的死亡年龄,可以计算得到死亡季节[108,130];④ 动物的一些部位终生生长并且受到季节变化的影响表现出周期性生长特征,通过观察生长纹可以得知动物的死亡季节,适合研究的材料包括贝壳、鱼类脊椎、鳃盖骨、耳石和哺乳动物牙骨质等[104];⑤通过食草类动物牙齿釉质氧同位素变化判断死亡季节[131].通过以上方法分析每个动物个体的死亡季节,可以得知一个物种的死亡季节分布;通过汇总所有物种所有个体的死亡季节,便可以得到遗址的使用季节.另外由于遗址堆积形成过程中受到多种埋藏学因素的影响,在利用动物遗存讨论获取季节时应尽量排除因自然营力进入遗址堆积的物种,选择明确因人类活动进入遗址堆积的样品,才能对遗址使用的季节性有更为全面、客观的了解[1,43,104]. ...
青藏高原的自然环境特征
1
2017
... 青藏高原平均海拔约4320 m,总面积308.34×104 km2 [44],被称为“世界屋脊”.高原地域环境特征鲜明,主要表现为高海拔、低温度、强辐射、河湖众多、冰川冻土广布、生物多样性丰富、高原内部自然环境差异显著[45].缺氧和高海拔造成的低生物生产能力,提高了捕获成本,且不能满足人类更大的营养需求;人类较低的生理能力,如低生育率和高死亡率,严重制约了人类早期从游猎高原到实现永久定居的发展历程[46].对史前人类定居青藏高原过程的研究,是探究人类对高海拔地区生理适应和行为适应的重要组成部分[47].动物作为肉食资源提供的高蛋白和高油脂,以及同时提供的毛、奶制品和畜力等二次产品,为人类适应高寒缺氧环境下的高能量消耗生活奠定了物质基础,因此,通过探讨早期先民的动物资源利用策略,可以揭示人类早期拓殖和定居高原的驱动机制.近年来,对史前人类在青藏高原的定居过程研究关注较多,但争议也多.本文系统梳理高原史前人类定居与动物资源利用的研究进展,并对利用动物遗存研究史前人类何时、如何定居青藏高提出因地制宜的方法和建议. ...
青藏高原的自然环境特征
1
2017
... 青藏高原平均海拔约4320 m,总面积308.34×104 km2 [44],被称为“世界屋脊”.高原地域环境特征鲜明,主要表现为高海拔、低温度、强辐射、河湖众多、冰川冻土广布、生物多样性丰富、高原内部自然环境差异显著[45].缺氧和高海拔造成的低生物生产能力,提高了捕获成本,且不能满足人类更大的营养需求;人类较低的生理能力,如低生育率和高死亡率,严重制约了人类早期从游猎高原到实现永久定居的发展历程[46].对史前人类定居青藏高原过程的研究,是探究人类对高海拔地区生理适应和行为适应的重要组成部分[47].动物作为肉食资源提供的高蛋白和高油脂,以及同时提供的毛、奶制品和畜力等二次产品,为人类适应高寒缺氧环境下的高能量消耗生活奠定了物质基础,因此,通过探讨早期先民的动物资源利用策略,可以揭示人类早期拓殖和定居高原的驱动机制.近年来,对史前人类在青藏高原的定居过程研究关注较多,但争议也多.本文系统梳理高原史前人类定居与动物资源利用的研究进展,并对利用动物遗存研究史前人类何时、如何定居青藏高提出因地制宜的方法和建议. ...
Early human occupation of the Tibetan Plateau
1
2018
... 青藏高原平均海拔约4320 m,总面积308.34×104 km2 [44],被称为“世界屋脊”.高原地域环境特征鲜明,主要表现为高海拔、低温度、强辐射、河湖众多、冰川冻土广布、生物多样性丰富、高原内部自然环境差异显著[45].缺氧和高海拔造成的低生物生产能力,提高了捕获成本,且不能满足人类更大的营养需求;人类较低的生理能力,如低生育率和高死亡率,严重制约了人类早期从游猎高原到实现永久定居的发展历程[46].对史前人类定居青藏高原过程的研究,是探究人类对高海拔地区生理适应和行为适应的重要组成部分[47].动物作为肉食资源提供的高蛋白和高油脂,以及同时提供的毛、奶制品和畜力等二次产品,为人类适应高寒缺氧环境下的高能量消耗生活奠定了物质基础,因此,通过探讨早期先民的动物资源利用策略,可以揭示人类早期拓殖和定居高原的驱动机制.近年来,对史前人类在青藏高原的定居过程研究关注较多,但争议也多.本文系统梳理高原史前人类定居与动物资源利用的研究进展,并对利用动物遗存研究史前人类何时、如何定居青藏高提出因地制宜的方法和建议. ...
History and possible mechanisms of prehistoric human migration to the Tibetan Plateau
3
2016
... 青藏高原平均海拔约4320 m,总面积308.34×104 km2 [44],被称为“世界屋脊”.高原地域环境特征鲜明,主要表现为高海拔、低温度、强辐射、河湖众多、冰川冻土广布、生物多样性丰富、高原内部自然环境差异显著[45].缺氧和高海拔造成的低生物生产能力,提高了捕获成本,且不能满足人类更大的营养需求;人类较低的生理能力,如低生育率和高死亡率,严重制约了人类早期从游猎高原到实现永久定居的发展历程[46].对史前人类定居青藏高原过程的研究,是探究人类对高海拔地区生理适应和行为适应的重要组成部分[47].动物作为肉食资源提供的高蛋白和高油脂,以及同时提供的毛、奶制品和畜力等二次产品,为人类适应高寒缺氧环境下的高能量消耗生活奠定了物质基础,因此,通过探讨早期先民的动物资源利用策略,可以揭示人类早期拓殖和定居高原的驱动机制.近年来,对史前人类在青藏高原的定居过程研究关注较多,但争议也多.本文系统梳理高原史前人类定居与动物资源利用的研究进展,并对利用动物遗存研究史前人类何时、如何定居青藏高提出因地制宜的方法和建议. ...
... 关于早期人群在青藏高原上开始营定居生活的时间、地点和驱动机制存在不同认识.考古学家根据青海同德宗日遗址和西藏昌都卡若遗址的房址、墓葬、农具、动植物遗存等证据,认为早在5.0 ka BP前后,人类已经定居在海拔超过3000 m的青藏高原东缘地带[68,86];张东菊等认为粟作农业的传播推动史前人类在6.0—4.0 ka BP进入青藏高原东北和东南部的低海拔河谷地带并定居,而后从西方传入的麦作农业促使了人群在3.6 ka BP向更高海拔常年定居[47];同时,根据高原东北部3.6—2.3 ka BP 5个诺木洪文化遗址的动植物考古学证据,学者提出放牧羊和牦牛及大麦种植为主的农牧并重的生产方式,支撑了海拔超过2800 m的诺木洪文化定居的形成[87].最近,陈发虎等将高原定居过程整理出3个阶段,即粟作农业人群至少在5.2 ka BP前进入高原东部低海拔的河谷地带,在4.8 ka BP以后开始定居在高原东部3000 m以上的高海拔区域;3.5 ka BP以来麦作的传入和牧业经济的发展助力人群大规模定居高海拔区域[87].但上述研究,除经过系统发掘的宗日、卡若和小恩达遗址,其他涉及的3000 m以上高海拔考古遗址目前还都缺少一年四季常年定居的直接证据. ...
... 以上关于史前人类全年游猎和定居青藏高原的理论大多建立在低地农业人群向高地输出的假设之上,农业的传入自然成为人类可以常年生活在青藏高原高海拔地区的前提.但“人类何时定居高原”和“青藏高原最早的农业”这两个问题是否能等同化[88]?非农业社会是否能够永久定居青藏高原?在青藏高原长久存在的高原狩猎采集者在早期定居和作物传播过程中扮演何种角色[47]?学者们认为,在讨论人类何时定居高原这一问题时,野生动物资源利用的作用被低估,低地河谷农业人群和高原狩猎采集人群之间的关系被忽略[88]. ...
A late middle Pleistocene Denisovan mandible from the Tibetan Plateau
2
2019
... 近些年的考古发现和发掘表明,早在200 ka BP,古老型智人丹尼索瓦人就已经进入青藏高原并长期活动,还可能在高原腹地留下岩面艺术[48⇓⇓-51];新近发掘的川西皮洛遗址(早于130 ka BP、海拔3750 m)出土了阿舍利手斧[52].现代智人(Homo sapiens)也于40—30 ka BP到达高原腹地海拔4480 m的尼阿底遗址(40—30 ka BP)[53],其在高原上一直游猎到全新世中晚期,151(15.4—13.1 ka BP、海拔3397 m)[54]、江西沟#1(14.6 ka BP、海拔3200 m)和西大滩#2(9.2—6.4 ka BP、海拔4569 m)[55]等遗址(图1)的发现,揭示了史前人类对青藏高原高海拔环境长久的适应历史. ...
... 注:1. 邱桑(226—169 ka BP、海拔4369 m)[50];2. 白石崖溶洞(190—45 ka BP、海拔3200 m)[48-49];3. 尼阿底;4. 151;5. 江西沟#1;6. 黑马河#1(13.1 ka BP、海拔3200 m)[56];7. 下大武(~11 ka BP、海拔3988 m)[57];8. 西大滩#2;9. 沙隆卡(8.3—8 ka BP、海拔2046 m)[58];10. 羊场(7.6—7.0 ka BP、海拔2440 m a.s.l.)[59];11. 野牛沟(7.5 ka BP、海拔3800 m)[60];12. 拉乙亥(~7.6 ka BP、海拔3125 m)[61];13. 仲巴10-1(6.6—2.6 ka BP、海拔4570 m)[62];14. 奖俊埠01(120—90 ka BP、海拔2673 m)[63];15. 皮洛. ...
Denisovan DNA in late Pleistocene sediments from Baishiya Karst Cave on the Tibetan plateau
4
2020
... 近些年的考古发现和发掘表明,早在200 ka BP,古老型智人丹尼索瓦人就已经进入青藏高原并长期活动,还可能在高原腹地留下岩面艺术[48⇓⇓-51];新近发掘的川西皮洛遗址(早于130 ka BP、海拔3750 m)出土了阿舍利手斧[52].现代智人(Homo sapiens)也于40—30 ka BP到达高原腹地海拔4480 m的尼阿底遗址(40—30 ka BP)[53],其在高原上一直游猎到全新世中晚期,151(15.4—13.1 ka BP、海拔3397 m)[54]、江西沟#1(14.6 ka BP、海拔3200 m)和西大滩#2(9.2—6.4 ka BP、海拔4569 m)[55]等遗址(图1)的发现,揭示了史前人类对青藏高原高海拔环境长久的适应历史. ...
... 注:1. 邱桑(226—169 ka BP、海拔4369 m)[50];2. 白石崖溶洞(190—45 ka BP、海拔3200 m)[48-49];3. 尼阿底;4. 151;5. 江西沟#1;6. 黑马河#1(13.1 ka BP、海拔3200 m)[56];7. 下大武(~11 ka BP、海拔3988 m)[57];8. 西大滩#2;9. 沙隆卡(8.3—8 ka BP、海拔2046 m)[58];10. 羊场(7.6—7.0 ka BP、海拔2440 m a.s.l.)[59];11. 野牛沟(7.5 ka BP、海拔3800 m)[60];12. 拉乙亥(~7.6 ka BP、海拔3125 m)[61];13. 仲巴10-1(6.6—2.6 ka BP、海拔4570 m)[62];14. 奖俊埠01(120—90 ka BP、海拔2673 m)[63];15. 皮洛. ...
... 青藏高原的旧石器时代遗址,除白石崖溶洞遗址(225—45 ka BP、海拔3200 m)[49]外,其他多数为旷野遗址,文化层较薄,揭露的灰坑、火塘等遗迹规模较小,出土遗物数量有限[54,57⇓⇓ -60,89 -90].通常认为,在此阶段人们以在高海拔和低海拔地区长距离移动的方式,间断性访问青藏高原高海拔地区[81,91],动物资源利用方式为纯粹的狩猎经济(图2).从利用的动物种类来看,旧石器中晚期(225—13.1 ka BP)以犀牛(Dicerorhinus)、马科(Equidae)、鹿科(Cervidae)、牛科(Bovidae)等大中型哺乳动物为主,在全新世早中期(10.4—5 ka BP)转变为以羊亚科(Caprinae)/羚羊亚科(Antilopinae)、啮齿类(Rodentia)和狐狸(Vulpes sp.)等中小型动物为主[49,54,56,61]. ...
... [49,54,56,61]. ...
Earliest parietal art: Hominin hand and foot traces from the middle Pleistocene of Tibet
2
2021
... 近些年的考古发现和发掘表明,早在200 ka BP,古老型智人丹尼索瓦人就已经进入青藏高原并长期活动,还可能在高原腹地留下岩面艺术[48⇓⇓-51];新近发掘的川西皮洛遗址(早于130 ka BP、海拔3750 m)出土了阿舍利手斧[52].现代智人(Homo sapiens)也于40—30 ka BP到达高原腹地海拔4480 m的尼阿底遗址(40—30 ka BP)[53],其在高原上一直游猎到全新世中晚期,151(15.4—13.1 ka BP、海拔3397 m)[54]、江西沟#1(14.6 ka BP、海拔3200 m)和西大滩#2(9.2—6.4 ka BP、海拔4569 m)[55]等遗址(图1)的发现,揭示了史前人类对青藏高原高海拔环境长久的适应历史. ...
... 注:1. 邱桑(226—169 ka BP、海拔4369 m)[50];2. 白石崖溶洞(190—45 ka BP、海拔3200 m)[48-49];3. 尼阿底;4. 151;5. 江西沟#1;6. 黑马河#1(13.1 ka BP、海拔3200 m)[56];7. 下大武(~11 ka BP、海拔3988 m)[57];8. 西大滩#2;9. 沙隆卡(8.3—8 ka BP、海拔2046 m)[58];10. 羊场(7.6—7.0 ka BP、海拔2440 m a.s.l.)[59];11. 野牛沟(7.5 ka BP、海拔3800 m)[60];12. 拉乙亥(~7.6 ka BP、海拔3125 m)[61];13. 仲巴10-1(6.6—2.6 ka BP、海拔4570 m)[62];14. 奖俊埠01(120—90 ka BP、海拔2673 m)[63];15. 皮洛. ...
Earliest hand and footprint art indicates that Denisovans may have occupied the interior of the high-altitude Tibetan Plateau since 200 thousand years ago
1
2022
... 近些年的考古发现和发掘表明,早在200 ka BP,古老型智人丹尼索瓦人就已经进入青藏高原并长期活动,还可能在高原腹地留下岩面艺术[48⇓⇓-51];新近发掘的川西皮洛遗址(早于130 ka BP、海拔3750 m)出土了阿舍利手斧[52].现代智人(Homo sapiens)也于40—30 ka BP到达高原腹地海拔4480 m的尼阿底遗址(40—30 ka BP)[53],其在高原上一直游猎到全新世中晚期,151(15.4—13.1 ka BP、海拔3397 m)[54]、江西沟#1(14.6 ka BP、海拔3200 m)和西大滩#2(9.2—6.4 ka BP、海拔4569 m)[55]等遗址(图1)的发现,揭示了史前人类对青藏高原高海拔环境长久的适应历史. ...
Denisovans and Homo sapiens on the Tibetan Plateau: Dispersals and adaptations
1
2022
... 近些年的考古发现和发掘表明,早在200 ka BP,古老型智人丹尼索瓦人就已经进入青藏高原并长期活动,还可能在高原腹地留下岩面艺术[48⇓⇓-51];新近发掘的川西皮洛遗址(早于130 ka BP、海拔3750 m)出土了阿舍利手斧[52].现代智人(Homo sapiens)也于40—30 ka BP到达高原腹地海拔4480 m的尼阿底遗址(40—30 ka BP)[53],其在高原上一直游猎到全新世中晚期,151(15.4—13.1 ka BP、海拔3397 m)[54]、江西沟#1(14.6 ka BP、海拔3200 m)和西大滩#2(9.2—6.4 ka BP、海拔4569 m)[55]等遗址(图1)的发现,揭示了史前人类对青藏高原高海拔环境长久的适应历史. ...
The earliest human occupation of the high-altitude Tibetan Plateau 40 thousand to 30 thousand years ago
1
2018
... 近些年的考古发现和发掘表明,早在200 ka BP,古老型智人丹尼索瓦人就已经进入青藏高原并长期活动,还可能在高原腹地留下岩面艺术[48⇓⇓-51];新近发掘的川西皮洛遗址(早于130 ka BP、海拔3750 m)出土了阿舍利手斧[52].现代智人(Homo sapiens)也于40—30 ka BP到达高原腹地海拔4480 m的尼阿底遗址(40—30 ka BP)[53],其在高原上一直游猎到全新世中晚期,151(15.4—13.1 ka BP、海拔3397 m)[54]、江西沟#1(14.6 ka BP、海拔3200 m)和西大滩#2(9.2—6.4 ka BP、海拔4569 m)[55]等遗址(图1)的发现,揭示了史前人类对青藏高原高海拔环境长久的适应历史. ...
Subsistence strategies of prehistoric hunter-gatherers on the Tibetan Plateau during the Last Deglaciation
3
2020
... 近些年的考古发现和发掘表明,早在200 ka BP,古老型智人丹尼索瓦人就已经进入青藏高原并长期活动,还可能在高原腹地留下岩面艺术[48⇓⇓-51];新近发掘的川西皮洛遗址(早于130 ka BP、海拔3750 m)出土了阿舍利手斧[52].现代智人(Homo sapiens)也于40—30 ka BP到达高原腹地海拔4480 m的尼阿底遗址(40—30 ka BP)[53],其在高原上一直游猎到全新世中晚期,151(15.4—13.1 ka BP、海拔3397 m)[54]、江西沟#1(14.6 ka BP、海拔3200 m)和西大滩#2(9.2—6.4 ka BP、海拔4569 m)[55]等遗址(图1)的发现,揭示了史前人类对青藏高原高海拔环境长久的适应历史. ...
... 青藏高原的旧石器时代遗址,除白石崖溶洞遗址(225—45 ka BP、海拔3200 m)[49]外,其他多数为旷野遗址,文化层较薄,揭露的灰坑、火塘等遗迹规模较小,出土遗物数量有限[54,57⇓⇓ -60,89 -90].通常认为,在此阶段人们以在高海拔和低海拔地区长距离移动的方式,间断性访问青藏高原高海拔地区[81,91],动物资源利用方式为纯粹的狩猎经济(图2).从利用的动物种类来看,旧石器中晚期(225—13.1 ka BP)以犀牛(Dicerorhinus)、马科(Equidae)、鹿科(Cervidae)、牛科(Bovidae)等大中型哺乳动物为主,在全新世早中期(10.4—5 ka BP)转变为以羊亚科(Caprinae)/羚羊亚科(Antilopinae)、啮齿类(Rodentia)和狐狸(Vulpes sp.)等中小型动物为主[49,54,56,61]. ...
... ,54,56,61]. ...
Late occupation of the high‐elevation northern Tibetan Plateau based on cosmogenic, luminescence, and radiocarbon ages
1
2013
... 近些年的考古发现和发掘表明,早在200 ka BP,古老型智人丹尼索瓦人就已经进入青藏高原并长期活动,还可能在高原腹地留下岩面艺术[48⇓⇓-51];新近发掘的川西皮洛遗址(早于130 ka BP、海拔3750 m)出土了阿舍利手斧[52].现代智人(Homo sapiens)也于40—30 ka BP到达高原腹地海拔4480 m的尼阿底遗址(40—30 ka BP)[53],其在高原上一直游猎到全新世中晚期,151(15.4—13.1 ka BP、海拔3397 m)[54]、江西沟#1(14.6 ka BP、海拔3200 m)和西大滩#2(9.2—6.4 ka BP、海拔4569 m)[55]等遗址(图1)的发现,揭示了史前人类对青藏高原高海拔环境长久的适应历史. ...
Peopling of the northern Tibetan Plateau
2
2006
... 注:1. 邱桑(226—169 ka BP、海拔4369 m)[50];2. 白石崖溶洞(190—45 ka BP、海拔3200 m)[48-49];3. 尼阿底;4. 151;5. 江西沟#1;6. 黑马河#1(13.1 ka BP、海拔3200 m)[56];7. 下大武(~11 ka BP、海拔3988 m)[57];8. 西大滩#2;9. 沙隆卡(8.3—8 ka BP、海拔2046 m)[58];10. 羊场(7.6—7.0 ka BP、海拔2440 m a.s.l.)[59];11. 野牛沟(7.5 ka BP、海拔3800 m)[60];12. 拉乙亥(~7.6 ka BP、海拔3125 m)[61];13. 仲巴10-1(6.6—2.6 ka BP、海拔4570 m)[62];14. 奖俊埠01(120—90 ka BP、海拔2673 m)[63];15. 皮洛. ...
... 青藏高原的旧石器时代遗址,除白石崖溶洞遗址(225—45 ka BP、海拔3200 m)[49]外,其他多数为旷野遗址,文化层较薄,揭露的灰坑、火塘等遗迹规模较小,出土遗物数量有限[54,57⇓⇓ -60,89 -90].通常认为,在此阶段人们以在高海拔和低海拔地区长距离移动的方式,间断性访问青藏高原高海拔地区[81,91],动物资源利用方式为纯粹的狩猎经济(图2).从利用的动物种类来看,旧石器中晚期(225—13.1 ka BP)以犀牛(Dicerorhinus)、马科(Equidae)、鹿科(Cervidae)、牛科(Bovidae)等大中型哺乳动物为主,在全新世早中期(10.4—5 ka BP)转变为以羊亚科(Caprinae)/羚羊亚科(Antilopinae)、啮齿类(Rodentia)和狐狸(Vulpes sp.)等中小型动物为主[49,54,56,61]. ...
青藏高原海拔4000 m区域人类活动的新证据
2
2016
... 注:1. 邱桑(226—169 ka BP、海拔4369 m)[50];2. 白石崖溶洞(190—45 ka BP、海拔3200 m)[48-49];3. 尼阿底;4. 151;5. 江西沟#1;6. 黑马河#1(13.1 ka BP、海拔3200 m)[56];7. 下大武(~11 ka BP、海拔3988 m)[57];8. 西大滩#2;9. 沙隆卡(8.3—8 ka BP、海拔2046 m)[58];10. 羊场(7.6—7.0 ka BP、海拔2440 m a.s.l.)[59];11. 野牛沟(7.5 ka BP、海拔3800 m)[60];12. 拉乙亥(~7.6 ka BP、海拔3125 m)[61];13. 仲巴10-1(6.6—2.6 ka BP、海拔4570 m)[62];14. 奖俊埠01(120—90 ka BP、海拔2673 m)[63];15. 皮洛. ...
... 青藏高原的旧石器时代遗址,除白石崖溶洞遗址(225—45 ka BP、海拔3200 m)[49]外,其他多数为旷野遗址,文化层较薄,揭露的灰坑、火塘等遗迹规模较小,出土遗物数量有限[54,57⇓⇓ -60,89 -90].通常认为,在此阶段人们以在高海拔和低海拔地区长距离移动的方式,间断性访问青藏高原高海拔地区[81,91],动物资源利用方式为纯粹的狩猎经济(图2).从利用的动物种类来看,旧石器中晚期(225—13.1 ka BP)以犀牛(Dicerorhinus)、马科(Equidae)、鹿科(Cervidae)、牛科(Bovidae)等大中型哺乳动物为主,在全新世早中期(10.4—5 ka BP)转变为以羊亚科(Caprinae)/羚羊亚科(Antilopinae)、啮齿类(Rodentia)和狐狸(Vulpes sp.)等中小型动物为主[49,54,56,61]. ...
青藏高原海拔4000 m区域人类活动的新证据
2
2016
... 注:1. 邱桑(226—169 ka BP、海拔4369 m)[50];2. 白石崖溶洞(190—45 ka BP、海拔3200 m)[48-49];3. 尼阿底;4. 151;5. 江西沟#1;6. 黑马河#1(13.1 ka BP、海拔3200 m)[56];7. 下大武(~11 ka BP、海拔3988 m)[57];8. 西大滩#2;9. 沙隆卡(8.3—8 ka BP、海拔2046 m)[58];10. 羊场(7.6—7.0 ka BP、海拔2440 m a.s.l.)[59];11. 野牛沟(7.5 ka BP、海拔3800 m)[60];12. 拉乙亥(~7.6 ka BP、海拔3125 m)[61];13. 仲巴10-1(6.6—2.6 ka BP、海拔4570 m)[62];14. 奖俊埠01(120—90 ka BP、海拔2673 m)[63];15. 皮洛. ...
... 青藏高原的旧石器时代遗址,除白石崖溶洞遗址(225—45 ka BP、海拔3200 m)[49]外,其他多数为旷野遗址,文化层较薄,揭露的灰坑、火塘等遗迹规模较小,出土遗物数量有限[54,57⇓⇓ -60,89 -90].通常认为,在此阶段人们以在高海拔和低海拔地区长距离移动的方式,间断性访问青藏高原高海拔地区[81,91],动物资源利用方式为纯粹的狩猎经济(图2).从利用的动物种类来看,旧石器中晚期(225—13.1 ka BP)以犀牛(Dicerorhinus)、马科(Equidae)、鹿科(Cervidae)、牛科(Bovidae)等大中型哺乳动物为主,在全新世早中期(10.4—5 ka BP)转变为以羊亚科(Caprinae)/羚羊亚科(Antilopinae)、啮齿类(Rodentia)和狐狸(Vulpes sp.)等中小型动物为主[49,54,56,61]. ...
Spatial and temporal variety of prehistoric human settlement and its influencing factors in the upper Yellow River valley, Qinghai Province, China
2
2013
... 注:1. 邱桑(226—169 ka BP、海拔4369 m)[50];2. 白石崖溶洞(190—45 ka BP、海拔3200 m)[48-49];3. 尼阿底;4. 151;5. 江西沟#1;6. 黑马河#1(13.1 ka BP、海拔3200 m)[56];7. 下大武(~11 ka BP、海拔3988 m)[57];8. 西大滩#2;9. 沙隆卡(8.3—8 ka BP、海拔2046 m)[58];10. 羊场(7.6—7.0 ka BP、海拔2440 m a.s.l.)[59];11. 野牛沟(7.5 ka BP、海拔3800 m)[60];12. 拉乙亥(~7.6 ka BP、海拔3125 m)[61];13. 仲巴10-1(6.6—2.6 ka BP、海拔4570 m)[62];14. 奖俊埠01(120—90 ka BP、海拔2673 m)[63];15. 皮洛. ...
... 青藏高原的旧石器时代遗址,除白石崖溶洞遗址(225—45 ka BP、海拔3200 m)[49]外,其他多数为旷野遗址,文化层较薄,揭露的灰坑、火塘等遗迹规模较小,出土遗物数量有限[54,57⇓⇓ -60,89 -90].通常认为,在此阶段人们以在高海拔和低海拔地区长距离移动的方式,间断性访问青藏高原高海拔地区[81,91],动物资源利用方式为纯粹的狩猎经济(图2).从利用的动物种类来看,旧石器中晚期(225—13.1 ka BP)以犀牛(Dicerorhinus)、马科(Equidae)、鹿科(Cervidae)、牛科(Bovidae)等大中型哺乳动物为主,在全新世早中期(10.4—5 ka BP)转变为以羊亚科(Caprinae)/羚羊亚科(Antilopinae)、啮齿类(Rodentia)和狐狸(Vulpes sp.)等中小型动物为主[49,54,56,61]. ...
The earliest well-dated archeological site in the hyper-arid Tarim Basin and its implications for prehistoric human migration and climatic change
2
2014
... 注:1. 邱桑(226—169 ka BP、海拔4369 m)[50];2. 白石崖溶洞(190—45 ka BP、海拔3200 m)[48-49];3. 尼阿底;4. 151;5. 江西沟#1;6. 黑马河#1(13.1 ka BP、海拔3200 m)[56];7. 下大武(~11 ka BP、海拔3988 m)[57];8. 西大滩#2;9. 沙隆卡(8.3—8 ka BP、海拔2046 m)[58];10. 羊场(7.6—7.0 ka BP、海拔2440 m a.s.l.)[59];11. 野牛沟(7.5 ka BP、海拔3800 m)[60];12. 拉乙亥(~7.6 ka BP、海拔3125 m)[61];13. 仲巴10-1(6.6—2.6 ka BP、海拔4570 m)[62];14. 奖俊埠01(120—90 ka BP、海拔2673 m)[63];15. 皮洛. ...
... 青藏高原的旧石器时代遗址,除白石崖溶洞遗址(225—45 ka BP、海拔3200 m)[49]外,其他多数为旷野遗址,文化层较薄,揭露的灰坑、火塘等遗迹规模较小,出土遗物数量有限[54,57⇓⇓ -60,89 -90].通常认为,在此阶段人们以在高海拔和低海拔地区长距离移动的方式,间断性访问青藏高原高海拔地区[81,91],动物资源利用方式为纯粹的狩猎经济(图2).从利用的动物种类来看,旧石器中晚期(225—13.1 ka BP)以犀牛(Dicerorhinus)、马科(Equidae)、鹿科(Cervidae)、牛科(Bovidae)等大中型哺乳动物为主,在全新世早中期(10.4—5 ka BP)转变为以羊亚科(Caprinae)/羚羊亚科(Antilopinae)、啮齿类(Rodentia)和狐狸(Vulpes sp.)等中小型动物为主[49,54,56,61]. ...
青海昆仑山山口发现的细石器考古新材料
2
2013
... 注:1. 邱桑(226—169 ka BP、海拔4369 m)[50];2. 白石崖溶洞(190—45 ka BP、海拔3200 m)[48-49];3. 尼阿底;4. 151;5. 江西沟#1;6. 黑马河#1(13.1 ka BP、海拔3200 m)[56];7. 下大武(~11 ka BP、海拔3988 m)[57];8. 西大滩#2;9. 沙隆卡(8.3—8 ka BP、海拔2046 m)[58];10. 羊场(7.6—7.0 ka BP、海拔2440 m a.s.l.)[59];11. 野牛沟(7.5 ka BP、海拔3800 m)[60];12. 拉乙亥(~7.6 ka BP、海拔3125 m)[61];13. 仲巴10-1(6.6—2.6 ka BP、海拔4570 m)[62];14. 奖俊埠01(120—90 ka BP、海拔2673 m)[63];15. 皮洛. ...
... 青藏高原的旧石器时代遗址,除白石崖溶洞遗址(225—45 ka BP、海拔3200 m)[49]外,其他多数为旷野遗址,文化层较薄,揭露的灰坑、火塘等遗迹规模较小,出土遗物数量有限[54,57⇓⇓ -60,89 -90].通常认为,在此阶段人们以在高海拔和低海拔地区长距离移动的方式,间断性访问青藏高原高海拔地区[81,91],动物资源利用方式为纯粹的狩猎经济(图2).从利用的动物种类来看,旧石器中晚期(225—13.1 ka BP)以犀牛(Dicerorhinus)、马科(Equidae)、鹿科(Cervidae)、牛科(Bovidae)等大中型哺乳动物为主,在全新世早中期(10.4—5 ka BP)转变为以羊亚科(Caprinae)/羚羊亚科(Antilopinae)、啮齿类(Rodentia)和狐狸(Vulpes sp.)等中小型动物为主[49,54,56,61]. ...
青海昆仑山山口发现的细石器考古新材料
2
2013
... 注:1. 邱桑(226—169 ka BP、海拔4369 m)[50];2. 白石崖溶洞(190—45 ka BP、海拔3200 m)[48-49];3. 尼阿底;4. 151;5. 江西沟#1;6. 黑马河#1(13.1 ka BP、海拔3200 m)[56];7. 下大武(~11 ka BP、海拔3988 m)[57];8. 西大滩#2;9. 沙隆卡(8.3—8 ka BP、海拔2046 m)[58];10. 羊场(7.6—7.0 ka BP、海拔2440 m a.s.l.)[59];11. 野牛沟(7.5 ka BP、海拔3800 m)[60];12. 拉乙亥(~7.6 ka BP、海拔3125 m)[61];13. 仲巴10-1(6.6—2.6 ka BP、海拔4570 m)[62];14. 奖俊埠01(120—90 ka BP、海拔2673 m)[63];15. 皮洛. ...
... 青藏高原的旧石器时代遗址,除白石崖溶洞遗址(225—45 ka BP、海拔3200 m)[49]外,其他多数为旷野遗址,文化层较薄,揭露的灰坑、火塘等遗迹规模较小,出土遗物数量有限[54,57⇓⇓ -60,89 -90].通常认为,在此阶段人们以在高海拔和低海拔地区长距离移动的方式,间断性访问青藏高原高海拔地区[81,91],动物资源利用方式为纯粹的狩猎经济(图2).从利用的动物种类来看,旧石器中晚期(225—13.1 ka BP)以犀牛(Dicerorhinus)、马科(Equidae)、鹿科(Cervidae)、牛科(Bovidae)等大中型哺乳动物为主,在全新世早中期(10.4—5 ka BP)转变为以羊亚科(Caprinae)/羚羊亚科(Antilopinae)、啮齿类(Rodentia)和狐狸(Vulpes sp.)等中小型动物为主[49,54,56,61]. ...
黄河上游拉乙亥中石器时代遗址发掘报告
2
1983
... 注:1. 邱桑(226—169 ka BP、海拔4369 m)[50];2. 白石崖溶洞(190—45 ka BP、海拔3200 m)[48-49];3. 尼阿底;4. 151;5. 江西沟#1;6. 黑马河#1(13.1 ka BP、海拔3200 m)[56];7. 下大武(~11 ka BP、海拔3988 m)[57];8. 西大滩#2;9. 沙隆卡(8.3—8 ka BP、海拔2046 m)[58];10. 羊场(7.6—7.0 ka BP、海拔2440 m a.s.l.)[59];11. 野牛沟(7.5 ka BP、海拔3800 m)[60];12. 拉乙亥(~7.6 ka BP、海拔3125 m)[61];13. 仲巴10-1(6.6—2.6 ka BP、海拔4570 m)[62];14. 奖俊埠01(120—90 ka BP、海拔2673 m)[63];15. 皮洛. ...
... 青藏高原的旧石器时代遗址,除白石崖溶洞遗址(225—45 ka BP、海拔3200 m)[49]外,其他多数为旷野遗址,文化层较薄,揭露的灰坑、火塘等遗迹规模较小,出土遗物数量有限[54,57⇓⇓ -60,89 -90].通常认为,在此阶段人们以在高海拔和低海拔地区长距离移动的方式,间断性访问青藏高原高海拔地区[81,91],动物资源利用方式为纯粹的狩猎经济(图2).从利用的动物种类来看,旧石器中晚期(225—13.1 ka BP)以犀牛(Dicerorhinus)、马科(Equidae)、鹿科(Cervidae)、牛科(Bovidae)等大中型哺乳动物为主,在全新世早中期(10.4—5 ka BP)转变为以羊亚科(Caprinae)/羚羊亚科(Antilopinae)、啮齿类(Rodentia)和狐狸(Vulpes sp.)等中小型动物为主[49,54,56,61]. ...
黄河上游拉乙亥中石器时代遗址发掘报告
2
1983
... 注:1. 邱桑(226—169 ka BP、海拔4369 m)[50];2. 白石崖溶洞(190—45 ka BP、海拔3200 m)[48-49];3. 尼阿底;4. 151;5. 江西沟#1;6. 黑马河#1(13.1 ka BP、海拔3200 m)[56];7. 下大武(~11 ka BP、海拔3988 m)[57];8. 西大滩#2;9. 沙隆卡(8.3—8 ka BP、海拔2046 m)[58];10. 羊场(7.6—7.0 ka BP、海拔2440 m a.s.l.)[59];11. 野牛沟(7.5 ka BP、海拔3800 m)[60];12. 拉乙亥(~7.6 ka BP、海拔3125 m)[61];13. 仲巴10-1(6.6—2.6 ka BP、海拔4570 m)[62];14. 奖俊埠01(120—90 ka BP、海拔2673 m)[63];15. 皮洛. ...
... 青藏高原的旧石器时代遗址,除白石崖溶洞遗址(225—45 ka BP、海拔3200 m)[49]外,其他多数为旷野遗址,文化层较薄,揭露的灰坑、火塘等遗迹规模较小,出土遗物数量有限[54,57⇓⇓ -60,89 -90].通常认为,在此阶段人们以在高海拔和低海拔地区长距离移动的方式,间断性访问青藏高原高海拔地区[81,91],动物资源利用方式为纯粹的狩猎经济(图2).从利用的动物种类来看,旧石器中晚期(225—13.1 ka BP)以犀牛(Dicerorhinus)、马科(Equidae)、鹿科(Cervidae)、牛科(Bovidae)等大中型哺乳动物为主,在全新世早中期(10.4—5 ka BP)转变为以羊亚科(Caprinae)/羚羊亚科(Antilopinae)、啮齿类(Rodentia)和狐狸(Vulpes sp.)等中小型动物为主[49,54,56,61]. ...
Radiocarbon dating of interdune paleo‐wetland deposits to constrain the age of mid‐to‐late Holocene microlithic artifacts from the Zhongba site, southwestern Qinghai‐Tibet Plateau
1
2014
... 注:1. 邱桑(226—169 ka BP、海拔4369 m)[50];2. 白石崖溶洞(190—45 ka BP、海拔3200 m)[48-49];3. 尼阿底;4. 151;5. 江西沟#1;6. 黑马河#1(13.1 ka BP、海拔3200 m)[56];7. 下大武(~11 ka BP、海拔3988 m)[57];8. 西大滩#2;9. 沙隆卡(8.3—8 ka BP、海拔2046 m)[58];10. 羊场(7.6—7.0 ka BP、海拔2440 m a.s.l.)[59];11. 野牛沟(7.5 ka BP、海拔3800 m)[60];12. 拉乙亥(~7.6 ka BP、海拔3125 m)[61];13. 仲巴10-1(6.6—2.6 ka BP、海拔4570 m)[62];14. 奖俊埠01(120—90 ka BP、海拔2673 m)[63];15. 皮洛. ...
Hominin occupation of the Tibetan Plateau during the Last Interglacial Complex
1
2021
... 注:1. 邱桑(226—169 ka BP、海拔4369 m)[50];2. 白石崖溶洞(190—45 ka BP、海拔3200 m)[48-49];3. 尼阿底;4. 151;5. 江西沟#1;6. 黑马河#1(13.1 ka BP、海拔3200 m)[56];7. 下大武(~11 ka BP、海拔3988 m)[57];8. 西大滩#2;9. 沙隆卡(8.3—8 ka BP、海拔2046 m)[58];10. 羊场(7.6—7.0 ka BP、海拔2440 m a.s.l.)[59];11. 野牛沟(7.5 ka BP、海拔3800 m)[60];12. 拉乙亥(~7.6 ka BP、海拔3125 m)[61];13. 仲巴10-1(6.6—2.6 ka BP、海拔4570 m)[62];14. 奖俊埠01(120—90 ka BP、海拔2673 m)[63];15. 皮洛. ...
四川马尔康县哈休遗址2006年的试掘
1
2010
... 5500 a BP前后,人类在高原的活动方式开始从游猎向定居转变.定居遗址最早出现在高原东缘,从东北部至川西,代表性遗址有青海同德宗日(5.6—4.0 ka BP、海拔2820 m)、四川马尔康哈休(5.5—4.7 ka BP、海拔2840 m)[65]、茂县营盘山(5.3—4.6 ka BP、海拔1710 m)[66]、金川刘家寨遗址(5.3—4.7 ka BP、海拔2650 m)[67],以及西藏昌都的卡若(4.8—4.0 ka BP、海拔3233 m)和小恩达遗址(4.8—4.2 ka BP、海拔3200 m)[68-69].定居生活从边缘向腹地扩散,至公元前两千纪中期在拉萨市北部出现以曲贡遗址(3.750—3.0 ka BP、海拔3686 m)[70]为代表的定居聚落.这些遗址出土的遗迹遗物,尤其是房址等遗迹,展示了定居这种生活方式从高原周边向腹地扩展的时空特征. ...
四川马尔康县哈休遗址2006年的试掘
1
2010
... 5500 a BP前后,人类在高原的活动方式开始从游猎向定居转变.定居遗址最早出现在高原东缘,从东北部至川西,代表性遗址有青海同德宗日(5.6—4.0 ka BP、海拔2820 m)、四川马尔康哈休(5.5—4.7 ka BP、海拔2840 m)[65]、茂县营盘山(5.3—4.6 ka BP、海拔1710 m)[66]、金川刘家寨遗址(5.3—4.7 ka BP、海拔2650 m)[67],以及西藏昌都的卡若(4.8—4.0 ka BP、海拔3233 m)和小恩达遗址(4.8—4.2 ka BP、海拔3200 m)[68-69].定居生活从边缘向腹地扩散,至公元前两千纪中期在拉萨市北部出现以曲贡遗址(3.750—3.0 ka BP、海拔3686 m)[70]为代表的定居聚落.这些遗址出土的遗迹遗物,尤其是房址等遗迹,展示了定居这种生活方式从高原周边向腹地扩展的时空特征. ...
1
2018
... 5500 a BP前后,人类在高原的活动方式开始从游猎向定居转变.定居遗址最早出现在高原东缘,从东北部至川西,代表性遗址有青海同德宗日(5.6—4.0 ka BP、海拔2820 m)、四川马尔康哈休(5.5—4.7 ka BP、海拔2840 m)[65]、茂县营盘山(5.3—4.6 ka BP、海拔1710 m)[66]、金川刘家寨遗址(5.3—4.7 ka BP、海拔2650 m)[67],以及西藏昌都的卡若(4.8—4.0 ka BP、海拔3233 m)和小恩达遗址(4.8—4.2 ka BP、海拔3200 m)[68-69].定居生活从边缘向腹地扩散,至公元前两千纪中期在拉萨市北部出现以曲贡遗址(3.750—3.0 ka BP、海拔3686 m)[70]为代表的定居聚落.这些遗址出土的遗迹遗物,尤其是房址等遗迹,展示了定居这种生活方式从高原周边向腹地扩展的时空特征. ...
1
2018
... 5500 a BP前后,人类在高原的活动方式开始从游猎向定居转变.定居遗址最早出现在高原东缘,从东北部至川西,代表性遗址有青海同德宗日(5.6—4.0 ka BP、海拔2820 m)、四川马尔康哈休(5.5—4.7 ka BP、海拔2840 m)[65]、茂县营盘山(5.3—4.6 ka BP、海拔1710 m)[66]、金川刘家寨遗址(5.3—4.7 ka BP、海拔2650 m)[67],以及西藏昌都的卡若(4.8—4.0 ka BP、海拔3233 m)和小恩达遗址(4.8—4.2 ka BP、海拔3200 m)[68-69].定居生活从边缘向腹地扩散,至公元前两千纪中期在拉萨市北部出现以曲贡遗址(3.750—3.0 ka BP、海拔3686 m)[70]为代表的定居聚落.这些遗址出土的遗迹遗物,尤其是房址等遗迹,展示了定居这种生活方式从高原周边向腹地扩展的时空特征. ...
四川金川县刘家寨遗址2011年发掘简报
1
2021
... 5500 a BP前后,人类在高原的活动方式开始从游猎向定居转变.定居遗址最早出现在高原东缘,从东北部至川西,代表性遗址有青海同德宗日(5.6—4.0 ka BP、海拔2820 m)、四川马尔康哈休(5.5—4.7 ka BP、海拔2840 m)[65]、茂县营盘山(5.3—4.6 ka BP、海拔1710 m)[66]、金川刘家寨遗址(5.3—4.7 ka BP、海拔2650 m)[67],以及西藏昌都的卡若(4.8—4.0 ka BP、海拔3233 m)和小恩达遗址(4.8—4.2 ka BP、海拔3200 m)[68-69].定居生活从边缘向腹地扩散,至公元前两千纪中期在拉萨市北部出现以曲贡遗址(3.750—3.0 ka BP、海拔3686 m)[70]为代表的定居聚落.这些遗址出土的遗迹遗物,尤其是房址等遗迹,展示了定居这种生活方式从高原周边向腹地扩展的时空特征. ...
四川金川县刘家寨遗址2011年发掘简报
1
2021
... 5500 a BP前后,人类在高原的活动方式开始从游猎向定居转变.定居遗址最早出现在高原东缘,从东北部至川西,代表性遗址有青海同德宗日(5.6—4.0 ka BP、海拔2820 m)、四川马尔康哈休(5.5—4.7 ka BP、海拔2840 m)[65]、茂县营盘山(5.3—4.6 ka BP、海拔1710 m)[66]、金川刘家寨遗址(5.3—4.7 ka BP、海拔2650 m)[67],以及西藏昌都的卡若(4.8—4.0 ka BP、海拔3233 m)和小恩达遗址(4.8—4.2 ka BP、海拔3200 m)[68-69].定居生活从边缘向腹地扩散,至公元前两千纪中期在拉萨市北部出现以曲贡遗址(3.750—3.0 ka BP、海拔3686 m)[70]为代表的定居聚落.这些遗址出土的遗迹遗物,尤其是房址等遗迹,展示了定居这种生活方式从高原周边向腹地扩展的时空特征. ...
2
1985
... 5500 a BP前后,人类在高原的活动方式开始从游猎向定居转变.定居遗址最早出现在高原东缘,从东北部至川西,代表性遗址有青海同德宗日(5.6—4.0 ka BP、海拔2820 m)、四川马尔康哈休(5.5—4.7 ka BP、海拔2840 m)[65]、茂县营盘山(5.3—4.6 ka BP、海拔1710 m)[66]、金川刘家寨遗址(5.3—4.7 ka BP、海拔2650 m)[67],以及西藏昌都的卡若(4.8—4.0 ka BP、海拔3233 m)和小恩达遗址(4.8—4.2 ka BP、海拔3200 m)[68-69].定居生活从边缘向腹地扩散,至公元前两千纪中期在拉萨市北部出现以曲贡遗址(3.750—3.0 ka BP、海拔3686 m)[70]为代表的定居聚落.这些遗址出土的遗迹遗物,尤其是房址等遗迹,展示了定居这种生活方式从高原周边向腹地扩展的时空特征. ...
... 关于早期人群在青藏高原上开始营定居生活的时间、地点和驱动机制存在不同认识.考古学家根据青海同德宗日遗址和西藏昌都卡若遗址的房址、墓葬、农具、动植物遗存等证据,认为早在5.0 ka BP前后,人类已经定居在海拔超过3000 m的青藏高原东缘地带[68,86];张东菊等认为粟作农业的传播推动史前人类在6.0—4.0 ka BP进入青藏高原东北和东南部的低海拔河谷地带并定居,而后从西方传入的麦作农业促使了人群在3.6 ka BP向更高海拔常年定居[47];同时,根据高原东北部3.6—2.3 ka BP 5个诺木洪文化遗址的动植物考古学证据,学者提出放牧羊和牦牛及大麦种植为主的农牧并重的生产方式,支撑了海拔超过2800 m的诺木洪文化定居的形成[87].最近,陈发虎等将高原定居过程整理出3个阶段,即粟作农业人群至少在5.2 ka BP前进入高原东部低海拔的河谷地带,在4.8 ka BP以后开始定居在高原东部3000 m以上的高海拔区域;3.5 ka BP以来麦作的传入和牧业经济的发展助力人群大规模定居高海拔区域[87].但上述研究,除经过系统发掘的宗日、卡若和小恩达遗址,其他涉及的3000 m以上高海拔考古遗址目前还都缺少一年四季常年定居的直接证据. ...
2
1985
... 5500 a BP前后,人类在高原的活动方式开始从游猎向定居转变.定居遗址最早出现在高原东缘,从东北部至川西,代表性遗址有青海同德宗日(5.6—4.0 ka BP、海拔2820 m)、四川马尔康哈休(5.5—4.7 ka BP、海拔2840 m)[65]、茂县营盘山(5.3—4.6 ka BP、海拔1710 m)[66]、金川刘家寨遗址(5.3—4.7 ka BP、海拔2650 m)[67],以及西藏昌都的卡若(4.8—4.0 ka BP、海拔3233 m)和小恩达遗址(4.8—4.2 ka BP、海拔3200 m)[68-69].定居生活从边缘向腹地扩散,至公元前两千纪中期在拉萨市北部出现以曲贡遗址(3.750—3.0 ka BP、海拔3686 m)[70]为代表的定居聚落.这些遗址出土的遗迹遗物,尤其是房址等遗迹,展示了定居这种生活方式从高原周边向腹地扩展的时空特征. ...
... 关于早期人群在青藏高原上开始营定居生活的时间、地点和驱动机制存在不同认识.考古学家根据青海同德宗日遗址和西藏昌都卡若遗址的房址、墓葬、农具、动植物遗存等证据,认为早在5.0 ka BP前后,人类已经定居在海拔超过3000 m的青藏高原东缘地带[68,86];张东菊等认为粟作农业的传播推动史前人类在6.0—4.0 ka BP进入青藏高原东北和东南部的低海拔河谷地带并定居,而后从西方传入的麦作农业促使了人群在3.6 ka BP向更高海拔常年定居[47];同时,根据高原东北部3.6—2.3 ka BP 5个诺木洪文化遗址的动植物考古学证据,学者提出放牧羊和牦牛及大麦种植为主的农牧并重的生产方式,支撑了海拔超过2800 m的诺木洪文化定居的形成[87].最近,陈发虎等将高原定居过程整理出3个阶段,即粟作农业人群至少在5.2 ka BP前进入高原东部低海拔的河谷地带,在4.8 ka BP以后开始定居在高原东部3000 m以上的高海拔区域;3.5 ka BP以来麦作的传入和牧业经济的发展助力人群大规模定居高海拔区域[87].但上述研究,除经过系统发掘的宗日、卡若和小恩达遗址,其他涉及的3000 m以上高海拔考古遗址目前还都缺少一年四季常年定居的直接证据. ...
西藏小恩达新石器时代遗址试掘简报
1
1990
... 5500 a BP前后,人类在高原的活动方式开始从游猎向定居转变.定居遗址最早出现在高原东缘,从东北部至川西,代表性遗址有青海同德宗日(5.6—4.0 ka BP、海拔2820 m)、四川马尔康哈休(5.5—4.7 ka BP、海拔2840 m)[65]、茂县营盘山(5.3—4.6 ka BP、海拔1710 m)[66]、金川刘家寨遗址(5.3—4.7 ka BP、海拔2650 m)[67],以及西藏昌都的卡若(4.8—4.0 ka BP、海拔3233 m)和小恩达遗址(4.8—4.2 ka BP、海拔3200 m)[68-69].定居生活从边缘向腹地扩散,至公元前两千纪中期在拉萨市北部出现以曲贡遗址(3.750—3.0 ka BP、海拔3686 m)[70]为代表的定居聚落.这些遗址出土的遗迹遗物,尤其是房址等遗迹,展示了定居这种生活方式从高原周边向腹地扩展的时空特征. ...
西藏小恩达新石器时代遗址试掘简报
1
1990
... 5500 a BP前后,人类在高原的活动方式开始从游猎向定居转变.定居遗址最早出现在高原东缘,从东北部至川西,代表性遗址有青海同德宗日(5.6—4.0 ka BP、海拔2820 m)、四川马尔康哈休(5.5—4.7 ka BP、海拔2840 m)[65]、茂县营盘山(5.3—4.6 ka BP、海拔1710 m)[66]、金川刘家寨遗址(5.3—4.7 ka BP、海拔2650 m)[67],以及西藏昌都的卡若(4.8—4.0 ka BP、海拔3233 m)和小恩达遗址(4.8—4.2 ka BP、海拔3200 m)[68-69].定居生活从边缘向腹地扩散,至公元前两千纪中期在拉萨市北部出现以曲贡遗址(3.750—3.0 ka BP、海拔3686 m)[70]为代表的定居聚落.这些遗址出土的遗迹遗物,尤其是房址等遗迹,展示了定居这种生活方式从高原周边向腹地扩展的时空特征. ...
2
1999
... 5500 a BP前后,人类在高原的活动方式开始从游猎向定居转变.定居遗址最早出现在高原东缘,从东北部至川西,代表性遗址有青海同德宗日(5.6—4.0 ka BP、海拔2820 m)、四川马尔康哈休(5.5—4.7 ka BP、海拔2840 m)[65]、茂县营盘山(5.3—4.6 ka BP、海拔1710 m)[66]、金川刘家寨遗址(5.3—4.7 ka BP、海拔2650 m)[67],以及西藏昌都的卡若(4.8—4.0 ka BP、海拔3233 m)和小恩达遗址(4.8—4.2 ka BP、海拔3200 m)[68-69].定居生活从边缘向腹地扩散,至公元前两千纪中期在拉萨市北部出现以曲贡遗址(3.750—3.0 ka BP、海拔3686 m)[70]为代表的定居聚落.这些遗址出土的遗迹遗物,尤其是房址等遗迹,展示了定居这种生活方式从高原周边向腹地扩展的时空特征. ...
... 在4.5—3.0 ka BP,史前东西方文化交流加强,驯化于西亚的黄牛、山羊(Capra hircus)和绵羊传入至青海省东北部和云南地区,青藏高原地区先民动物资源利用在空间上呈现多元化,牛和羊超过了猪,成为家畜饲养的主流(图2).家羊(绵羊/山羊)在青藏高原最早出土于金蝉口遗址(4.5—3.6 ka BP、海拔2419 m).长宁遗址(4.3—3.6 ka BP、海拔2300 m)出土的动物遗存以家畜黄牛、山羊和绵羊为主,牛、羊所占比均超过了猪,在先民生业经济中占据主要地位[42,95].青藏高原南部拉萨曲贡遗址先民的生业经济以作物种植和放牧牦牛、绵羊的农牧经济为主,以猎取白唇鹿或马鹿、麝、野猪、藏野驴和各种禽类的狩猎经济为辅[70].在高原西部阿里扎达的格布赛鲁遗址早期墓葬(3560—3000 a BP、海拔3800 m)中随葬有山羊、绵羊和黄牛等家养动物,以及盘羊和鹿等野生动物;遗址晚期墓葬(2310—2127 a BP)中出土了山羊、绵羊、黄牛、马、野驴和鱼等.在3.0 ka BP以后,绵羊、山羊、黄牛、马、狗和牦牛等家养动物在青藏高原被广泛畜养,并在人们日常生活和社会仪式中扮演重要角色[100⇓⇓-103](图2). ...
2
1999
... 5500 a BP前后,人类在高原的活动方式开始从游猎向定居转变.定居遗址最早出现在高原东缘,从东北部至川西,代表性遗址有青海同德宗日(5.6—4.0 ka BP、海拔2820 m)、四川马尔康哈休(5.5—4.7 ka BP、海拔2840 m)[65]、茂县营盘山(5.3—4.6 ka BP、海拔1710 m)[66]、金川刘家寨遗址(5.3—4.7 ka BP、海拔2650 m)[67],以及西藏昌都的卡若(4.8—4.0 ka BP、海拔3233 m)和小恩达遗址(4.8—4.2 ka BP、海拔3200 m)[68-69].定居生活从边缘向腹地扩散,至公元前两千纪中期在拉萨市北部出现以曲贡遗址(3.750—3.0 ka BP、海拔3686 m)[70]为代表的定居聚落.这些遗址出土的遗迹遗物,尤其是房址等遗迹,展示了定居这种生活方式从高原周边向腹地扩展的时空特征. ...
... 在4.5—3.0 ka BP,史前东西方文化交流加强,驯化于西亚的黄牛、山羊(Capra hircus)和绵羊传入至青海省东北部和云南地区,青藏高原地区先民动物资源利用在空间上呈现多元化,牛和羊超过了猪,成为家畜饲养的主流(图2).家羊(绵羊/山羊)在青藏高原最早出土于金蝉口遗址(4.5—3.6 ka BP、海拔2419 m).长宁遗址(4.3—3.6 ka BP、海拔2300 m)出土的动物遗存以家畜黄牛、山羊和绵羊为主,牛、羊所占比均超过了猪,在先民生业经济中占据主要地位[42,95].青藏高原南部拉萨曲贡遗址先民的生业经济以作物种植和放牧牦牛、绵羊的农牧经济为主,以猎取白唇鹿或马鹿、麝、野猪、藏野驴和各种禽类的狩猎经济为辅[70].在高原西部阿里扎达的格布赛鲁遗址早期墓葬(3560—3000 a BP、海拔3800 m)中随葬有山羊、绵羊和黄牛等家养动物,以及盘羊和鹿等野生动物;遗址晚期墓葬(2310—2127 a BP)中出土了山羊、绵羊、黄牛、马、野驴和鱼等.在3.0 ka BP以后,绵羊、山羊、黄牛、马、狗和牦牛等家养动物在青藏高原被广泛畜养,并在人们日常生活和社会仪式中扮演重要角色[100⇓⇓-103](图2). ...
Mitochondrial genome evidence reveals successful Late Paleolithic settlement on the Tibetan Plateau
1
2009
... 遗传学分析结果表明,现代藏族人中有少部分来自更新世旧石器时代人群(62—38 ka BP)的古老成分[71⇓-73],如源自于古老型智人——丹尼索瓦人的EPAS1基因[74].Huerta-Sánchez等[74]对EPAS1基因传入现代人和受到强烈自然选择的时间进行了模拟,发现可能在9 ka BP发生了人群大规模向高原扩散的事件.另外,全基因组研究结果表明,现代青藏高原人群大部遗传成分来自15—9 ka BP古代东亚北方低海拔人群的大规模扩散[73,75],并在全新世后期存在复杂的人群扩散事件.已有遗传研究结合考古发现认为在6.5—2.5 ka BP可能存在黄河流域农业人群分别向青藏高原东北缘和中国西南地区的两次人群扩张,其中约在3600 a BP发生了中国北方的粟作农民带着麦作农业向高原扩散[76-77].但对于是人群迁移还是文化传播仍存在争议,已有古DNA研究结果表明,青藏高原史前人群对现代高原人群只有部分基因贡献,一部分现代藏族的祖先可能为4750—2775 a BP生活在低海拔地区的古代人群,且可能在2125—1100 a BP存在高低海拔地区之间的人群交流[78].除此以外,青藏高原现代人群还存在着少量与中亚、欧亚草原和南亚人群的基因交流[73],这些人群内部差异是如何产生以及何时产生的,并且相互间有何影响,仍然难以解答.综上,虽然在更新世晚期高原人群遗传成分存在并延续,全新世以来人类定居高原的过程可能是高原遗传成分形成的重要阶段,尤其是在农业传播以前,依靠大型哺乳动物或渔猎资源可能已经形成了独特高原人群遗传成分,而后伴随农业传播以及可能的牦牛驯化,不同区域不同的生业方式逐渐促使高原人群内部产生遗传分异.然而由于样本所限,遗传学所揭示的高原人群迁徙研究结果还需要进一步明确,并需要和考古证据结合进行讨论. ...
Genetic evidence of paleolithic colonization and neolithic expansion of modern humans on the Tibetan Plateau
1
2013
... 遗传学分析结果表明,现代藏族人中有少部分来自更新世旧石器时代人群(62—38 ka BP)的古老成分[71⇓-73],如源自于古老型智人——丹尼索瓦人的EPAS1基因[74].Huerta-Sánchez等[74]对EPAS1基因传入现代人和受到强烈自然选择的时间进行了模拟,发现可能在9 ka BP发生了人群大规模向高原扩散的事件.另外,全基因组研究结果表明,现代青藏高原人群大部遗传成分来自15—9 ka BP古代东亚北方低海拔人群的大规模扩散[73,75],并在全新世后期存在复杂的人群扩散事件.已有遗传研究结合考古发现认为在6.5—2.5 ka BP可能存在黄河流域农业人群分别向青藏高原东北缘和中国西南地区的两次人群扩张,其中约在3600 a BP发生了中国北方的粟作农民带着麦作农业向高原扩散[76-77].但对于是人群迁移还是文化传播仍存在争议,已有古DNA研究结果表明,青藏高原史前人群对现代高原人群只有部分基因贡献,一部分现代藏族的祖先可能为4750—2775 a BP生活在低海拔地区的古代人群,且可能在2125—1100 a BP存在高低海拔地区之间的人群交流[78].除此以外,青藏高原现代人群还存在着少量与中亚、欧亚草原和南亚人群的基因交流[73],这些人群内部差异是如何产生以及何时产生的,并且相互间有何影响,仍然难以解答.综上,虽然在更新世晚期高原人群遗传成分存在并延续,全新世以来人类定居高原的过程可能是高原遗传成分形成的重要阶段,尤其是在农业传播以前,依靠大型哺乳动物或渔猎资源可能已经形成了独特高原人群遗传成分,而后伴随农业传播以及可能的牦牛驯化,不同区域不同的生业方式逐渐促使高原人群内部产生遗传分异.然而由于样本所限,遗传学所揭示的高原人群迁徙研究结果还需要进一步明确,并需要和考古证据结合进行讨论. ...
Ancestral origins and genetic history of Tibetan highlanders
3
2016
... 遗传学分析结果表明,现代藏族人中有少部分来自更新世旧石器时代人群(62—38 ka BP)的古老成分[71⇓-73],如源自于古老型智人——丹尼索瓦人的EPAS1基因[74].Huerta-Sánchez等[74]对EPAS1基因传入现代人和受到强烈自然选择的时间进行了模拟,发现可能在9 ka BP发生了人群大规模向高原扩散的事件.另外,全基因组研究结果表明,现代青藏高原人群大部遗传成分来自15—9 ka BP古代东亚北方低海拔人群的大规模扩散[73,75],并在全新世后期存在复杂的人群扩散事件.已有遗传研究结合考古发现认为在6.5—2.5 ka BP可能存在黄河流域农业人群分别向青藏高原东北缘和中国西南地区的两次人群扩张,其中约在3600 a BP发生了中国北方的粟作农民带着麦作农业向高原扩散[76-77].但对于是人群迁移还是文化传播仍存在争议,已有古DNA研究结果表明,青藏高原史前人群对现代高原人群只有部分基因贡献,一部分现代藏族的祖先可能为4750—2775 a BP生活在低海拔地区的古代人群,且可能在2125—1100 a BP存在高低海拔地区之间的人群交流[78].除此以外,青藏高原现代人群还存在着少量与中亚、欧亚草原和南亚人群的基因交流[73],这些人群内部差异是如何产生以及何时产生的,并且相互间有何影响,仍然难以解答.综上,虽然在更新世晚期高原人群遗传成分存在并延续,全新世以来人类定居高原的过程可能是高原遗传成分形成的重要阶段,尤其是在农业传播以前,依靠大型哺乳动物或渔猎资源可能已经形成了独特高原人群遗传成分,而后伴随农业传播以及可能的牦牛驯化,不同区域不同的生业方式逐渐促使高原人群内部产生遗传分异.然而由于样本所限,遗传学所揭示的高原人群迁徙研究结果还需要进一步明确,并需要和考古证据结合进行讨论. ...
... [73,75],并在全新世后期存在复杂的人群扩散事件.已有遗传研究结合考古发现认为在6.5—2.5 ka BP可能存在黄河流域农业人群分别向青藏高原东北缘和中国西南地区的两次人群扩张,其中约在3600 a BP发生了中国北方的粟作农民带着麦作农业向高原扩散[76-77].但对于是人群迁移还是文化传播仍存在争议,已有古DNA研究结果表明,青藏高原史前人群对现代高原人群只有部分基因贡献,一部分现代藏族的祖先可能为4750—2775 a BP生活在低海拔地区的古代人群,且可能在2125—1100 a BP存在高低海拔地区之间的人群交流[78].除此以外,青藏高原现代人群还存在着少量与中亚、欧亚草原和南亚人群的基因交流[73],这些人群内部差异是如何产生以及何时产生的,并且相互间有何影响,仍然难以解答.综上,虽然在更新世晚期高原人群遗传成分存在并延续,全新世以来人类定居高原的过程可能是高原遗传成分形成的重要阶段,尤其是在农业传播以前,依靠大型哺乳动物或渔猎资源可能已经形成了独特高原人群遗传成分,而后伴随农业传播以及可能的牦牛驯化,不同区域不同的生业方式逐渐促使高原人群内部产生遗传分异.然而由于样本所限,遗传学所揭示的高原人群迁徙研究结果还需要进一步明确,并需要和考古证据结合进行讨论. ...
... [73],这些人群内部差异是如何产生以及何时产生的,并且相互间有何影响,仍然难以解答.综上,虽然在更新世晚期高原人群遗传成分存在并延续,全新世以来人类定居高原的过程可能是高原遗传成分形成的重要阶段,尤其是在农业传播以前,依靠大型哺乳动物或渔猎资源可能已经形成了独特高原人群遗传成分,而后伴随农业传播以及可能的牦牛驯化,不同区域不同的生业方式逐渐促使高原人群内部产生遗传分异.然而由于样本所限,遗传学所揭示的高原人群迁徙研究结果还需要进一步明确,并需要和考古证据结合进行讨论. ...
Altitude adaptation in Tibetans caused by introgression of Denisovan-like DNA
2
2014
... 遗传学分析结果表明,现代藏族人中有少部分来自更新世旧石器时代人群(62—38 ka BP)的古老成分[71⇓-73],如源自于古老型智人——丹尼索瓦人的EPAS1基因[74].Huerta-Sánchez等[74]对EPAS1基因传入现代人和受到强烈自然选择的时间进行了模拟,发现可能在9 ka BP发生了人群大规模向高原扩散的事件.另外,全基因组研究结果表明,现代青藏高原人群大部遗传成分来自15—9 ka BP古代东亚北方低海拔人群的大规模扩散[73,75],并在全新世后期存在复杂的人群扩散事件.已有遗传研究结合考古发现认为在6.5—2.5 ka BP可能存在黄河流域农业人群分别向青藏高原东北缘和中国西南地区的两次人群扩张,其中约在3600 a BP发生了中国北方的粟作农民带着麦作农业向高原扩散[76-77].但对于是人群迁移还是文化传播仍存在争议,已有古DNA研究结果表明,青藏高原史前人群对现代高原人群只有部分基因贡献,一部分现代藏族的祖先可能为4750—2775 a BP生活在低海拔地区的古代人群,且可能在2125—1100 a BP存在高低海拔地区之间的人群交流[78].除此以外,青藏高原现代人群还存在着少量与中亚、欧亚草原和南亚人群的基因交流[73],这些人群内部差异是如何产生以及何时产生的,并且相互间有何影响,仍然难以解答.综上,虽然在更新世晚期高原人群遗传成分存在并延续,全新世以来人类定居高原的过程可能是高原遗传成分形成的重要阶段,尤其是在农业传播以前,依靠大型哺乳动物或渔猎资源可能已经形成了独特高原人群遗传成分,而后伴随农业传播以及可能的牦牛驯化,不同区域不同的生业方式逐渐促使高原人群内部产生遗传分异.然而由于样本所限,遗传学所揭示的高原人群迁徙研究结果还需要进一步明确,并需要和考古证据结合进行讨论. ...
... [74]对EPAS1基因传入现代人和受到强烈自然选择的时间进行了模拟,发现可能在9 ka BP发生了人群大规模向高原扩散的事件.另外,全基因组研究结果表明,现代青藏高原人群大部遗传成分来自15—9 ka BP古代东亚北方低海拔人群的大规模扩散[73,75],并在全新世后期存在复杂的人群扩散事件.已有遗传研究结合考古发现认为在6.5—2.5 ka BP可能存在黄河流域农业人群分别向青藏高原东北缘和中国西南地区的两次人群扩张,其中约在3600 a BP发生了中国北方的粟作农民带着麦作农业向高原扩散[76-77].但对于是人群迁移还是文化传播仍存在争议,已有古DNA研究结果表明,青藏高原史前人群对现代高原人群只有部分基因贡献,一部分现代藏族的祖先可能为4750—2775 a BP生活在低海拔地区的古代人群,且可能在2125—1100 a BP存在高低海拔地区之间的人群交流[78].除此以外,青藏高原现代人群还存在着少量与中亚、欧亚草原和南亚人群的基因交流[73],这些人群内部差异是如何产生以及何时产生的,并且相互间有何影响,仍然难以解答.综上,虽然在更新世晚期高原人群遗传成分存在并延续,全新世以来人类定居高原的过程可能是高原遗传成分形成的重要阶段,尤其是在农业传播以前,依靠大型哺乳动物或渔猎资源可能已经形成了独特高原人群遗传成分,而后伴随农业传播以及可能的牦牛驯化,不同区域不同的生业方式逐渐促使高原人群内部产生遗传分异.然而由于样本所限,遗传学所揭示的高原人群迁徙研究结果还需要进一步明确,并需要和考古证据结合进行讨论. ...
Evolutionary history of Tibetans inferred from whole-genome sequencing
1
2017
... 遗传学分析结果表明,现代藏族人中有少部分来自更新世旧石器时代人群(62—38 ka BP)的古老成分[71⇓-73],如源自于古老型智人——丹尼索瓦人的EPAS1基因[74].Huerta-Sánchez等[74]对EPAS1基因传入现代人和受到强烈自然选择的时间进行了模拟,发现可能在9 ka BP发生了人群大规模向高原扩散的事件.另外,全基因组研究结果表明,现代青藏高原人群大部遗传成分来自15—9 ka BP古代东亚北方低海拔人群的大规模扩散[73,75],并在全新世后期存在复杂的人群扩散事件.已有遗传研究结合考古发现认为在6.5—2.5 ka BP可能存在黄河流域农业人群分别向青藏高原东北缘和中国西南地区的两次人群扩张,其中约在3600 a BP发生了中国北方的粟作农民带着麦作农业向高原扩散[76-77].但对于是人群迁移还是文化传播仍存在争议,已有古DNA研究结果表明,青藏高原史前人群对现代高原人群只有部分基因贡献,一部分现代藏族的祖先可能为4750—2775 a BP生活在低海拔地区的古代人群,且可能在2125—1100 a BP存在高低海拔地区之间的人群交流[78].除此以外,青藏高原现代人群还存在着少量与中亚、欧亚草原和南亚人群的基因交流[73],这些人群内部差异是如何产生以及何时产生的,并且相互间有何影响,仍然难以解答.综上,虽然在更新世晚期高原人群遗传成分存在并延续,全新世以来人类定居高原的过程可能是高原遗传成分形成的重要阶段,尤其是在农业传播以前,依靠大型哺乳动物或渔猎资源可能已经形成了独特高原人群遗传成分,而后伴随农业传播以及可能的牦牛驯化,不同区域不同的生业方式逐渐促使高原人群内部产生遗传分异.然而由于样本所限,遗传学所揭示的高原人群迁徙研究结果还需要进一步明确,并需要和考古证据结合进行讨论. ...
Reconstruction of Y-chromosome phylogeny reveals two neolithic expansions of Tibeto-Burman populations
1
2018
... 遗传学分析结果表明,现代藏族人中有少部分来自更新世旧石器时代人群(62—38 ka BP)的古老成分[71⇓-73],如源自于古老型智人——丹尼索瓦人的EPAS1基因[74].Huerta-Sánchez等[74]对EPAS1基因传入现代人和受到强烈自然选择的时间进行了模拟,发现可能在9 ka BP发生了人群大规模向高原扩散的事件.另外,全基因组研究结果表明,现代青藏高原人群大部遗传成分来自15—9 ka BP古代东亚北方低海拔人群的大规模扩散[73,75],并在全新世后期存在复杂的人群扩散事件.已有遗传研究结合考古发现认为在6.5—2.5 ka BP可能存在黄河流域农业人群分别向青藏高原东北缘和中国西南地区的两次人群扩张,其中约在3600 a BP发生了中国北方的粟作农民带着麦作农业向高原扩散[76-77].但对于是人群迁移还是文化传播仍存在争议,已有古DNA研究结果表明,青藏高原史前人群对现代高原人群只有部分基因贡献,一部分现代藏族的祖先可能为4750—2775 a BP生活在低海拔地区的古代人群,且可能在2125—1100 a BP存在高低海拔地区之间的人群交流[78].除此以外,青藏高原现代人群还存在着少量与中亚、欧亚草原和南亚人群的基因交流[73],这些人群内部差异是如何产生以及何时产生的,并且相互间有何影响,仍然难以解答.综上,虽然在更新世晚期高原人群遗传成分存在并延续,全新世以来人类定居高原的过程可能是高原遗传成分形成的重要阶段,尤其是在农业传播以前,依靠大型哺乳动物或渔猎资源可能已经形成了独特高原人群遗传成分,而后伴随农业传播以及可能的牦牛驯化,不同区域不同的生业方式逐渐促使高原人群内部产生遗传分异.然而由于样本所限,遗传学所揭示的高原人群迁徙研究结果还需要进一步明确,并需要和考古证据结合进行讨论. ...
Neolithic millet farmers contributed to the permanent settlement of the Tibetan Plateau by adopting barley agriculture
1
2019
... 遗传学分析结果表明,现代藏族人中有少部分来自更新世旧石器时代人群(62—38 ka BP)的古老成分[71⇓-73],如源自于古老型智人——丹尼索瓦人的EPAS1基因[74].Huerta-Sánchez等[74]对EPAS1基因传入现代人和受到强烈自然选择的时间进行了模拟,发现可能在9 ka BP发生了人群大规模向高原扩散的事件.另外,全基因组研究结果表明,现代青藏高原人群大部遗传成分来自15—9 ka BP古代东亚北方低海拔人群的大规模扩散[73,75],并在全新世后期存在复杂的人群扩散事件.已有遗传研究结合考古发现认为在6.5—2.5 ka BP可能存在黄河流域农业人群分别向青藏高原东北缘和中国西南地区的两次人群扩张,其中约在3600 a BP发生了中国北方的粟作农民带着麦作农业向高原扩散[76-77].但对于是人群迁移还是文化传播仍存在争议,已有古DNA研究结果表明,青藏高原史前人群对现代高原人群只有部分基因贡献,一部分现代藏族的祖先可能为4750—2775 a BP生活在低海拔地区的古代人群,且可能在2125—1100 a BP存在高低海拔地区之间的人群交流[78].除此以外,青藏高原现代人群还存在着少量与中亚、欧亚草原和南亚人群的基因交流[73],这些人群内部差异是如何产生以及何时产生的,并且相互间有何影响,仍然难以解答.综上,虽然在更新世晚期高原人群遗传成分存在并延续,全新世以来人类定居高原的过程可能是高原遗传成分形成的重要阶段,尤其是在农业传播以前,依靠大型哺乳动物或渔猎资源可能已经形成了独特高原人群遗传成分,而后伴随农业传播以及可能的牦牛驯化,不同区域不同的生业方式逐渐促使高原人群内部产生遗传分异.然而由于样本所限,遗传学所揭示的高原人群迁徙研究结果还需要进一步明确,并需要和考古证据结合进行讨论. ...
Ancient mitogenomes show plateau populations from last 5200 years partially contributed to present-day Tibetans
1
1923
... 遗传学分析结果表明,现代藏族人中有少部分来自更新世旧石器时代人群(62—38 ka BP)的古老成分[71⇓-73],如源自于古老型智人——丹尼索瓦人的EPAS1基因[74].Huerta-Sánchez等[74]对EPAS1基因传入现代人和受到强烈自然选择的时间进行了模拟,发现可能在9 ka BP发生了人群大规模向高原扩散的事件.另外,全基因组研究结果表明,现代青藏高原人群大部遗传成分来自15—9 ka BP古代东亚北方低海拔人群的大规模扩散[73,75],并在全新世后期存在复杂的人群扩散事件.已有遗传研究结合考古发现认为在6.5—2.5 ka BP可能存在黄河流域农业人群分别向青藏高原东北缘和中国西南地区的两次人群扩张,其中约在3600 a BP发生了中国北方的粟作农民带着麦作农业向高原扩散[76-77].但对于是人群迁移还是文化传播仍存在争议,已有古DNA研究结果表明,青藏高原史前人群对现代高原人群只有部分基因贡献,一部分现代藏族的祖先可能为4750—2775 a BP生活在低海拔地区的古代人群,且可能在2125—1100 a BP存在高低海拔地区之间的人群交流[78].除此以外,青藏高原现代人群还存在着少量与中亚、欧亚草原和南亚人群的基因交流[73],这些人群内部差异是如何产生以及何时产生的,并且相互间有何影响,仍然难以解答.综上,虽然在更新世晚期高原人群遗传成分存在并延续,全新世以来人类定居高原的过程可能是高原遗传成分形成的重要阶段,尤其是在农业传播以前,依靠大型哺乳动物或渔猎资源可能已经形成了独特高原人群遗传成分,而后伴随农业传播以及可能的牦牛驯化,不同区域不同的生业方式逐渐促使高原人群内部产生遗传分异.然而由于样本所限,遗传学所揭示的高原人群迁徙研究结果还需要进一步明确,并需要和考古证据结合进行讨论. ...
史前时代人类向青藏高原扩散的过程与动力
1
2016
... 全年生活在青藏高原包括两个层面的意思,一是全年游猎于高原,一是全年固定于一处地方生活,即定居.一些研究中提及的“year round/permanent occupation”主要强调全年生活于高海拔地区[79-80],非本文讨论的定居,为避免行文引起歧义,本文用“占据”(occupation)一词.二者在逻辑上有一定联系,全年占据并生活在青藏高原的社会形态不一定是定居,但定居肯定包括在全年占据这一人类活动之中;在全年占据的基础上,需要对遗址进行进一步的发掘和研究,尤其是遗址使用的季节性分析——只有一年四季且常年使用,才属于本文所讨论的考古学意义上的定居. ...
史前时代人类向青藏高原扩散的过程与动力
1
2016
... 全年生活在青藏高原包括两个层面的意思,一是全年游猎于高原,一是全年固定于一处地方生活,即定居.一些研究中提及的“year round/permanent occupation”主要强调全年生活于高海拔地区[79-80],非本文讨论的定居,为避免行文引起歧义,本文用“占据”(occupation)一词.二者在逻辑上有一定联系,全年占据并生活在青藏高原的社会形态不一定是定居,但定居肯定包括在全年占据这一人类活动之中;在全年占据的基础上,需要对遗址进行进一步的发掘和研究,尤其是遗址使用的季节性分析——只有一年四季且常年使用,才属于本文所讨论的考古学意义上的定居. ...
Permanent human occupation of the central Tibetan Plateau in the early Holocene
1
2017
... 全年生活在青藏高原包括两个层面的意思,一是全年游猎于高原,一是全年固定于一处地方生活,即定居.一些研究中提及的“year round/permanent occupation”主要强调全年生活于高海拔地区[79-80],非本文讨论的定居,为避免行文引起歧义,本文用“占据”(occupation)一词.二者在逻辑上有一定联系,全年占据并生活在青藏高原的社会形态不一定是定居,但定居肯定包括在全年占据这一人类活动之中;在全年占据的基础上,需要对遗址进行进一步的发掘和研究,尤其是遗址使用的季节性分析——只有一年四季且常年使用,才属于本文所讨论的考古学意义上的定居. ...
Archaeology augments Tibet's genetic history
2
2010
... 部分学者认为人群全年占据青藏高原海拔3300 m以上地区的时间不早于7.0 ka BP,且与低海拔农业人群保持密切联系,而人群全年占据海拔4000 m以上地区不早于4.0 ka BP,且与牦牛(Bos grunniens)驯养关系密切[81].来自青藏高原东北部的动植物遗存的证据则表明,3600 a BP麦作农业的传入促进人类大规模占据青藏高原2500 m以上高海拔地区[82].另外,驯化动物,如绵羊(Ovis aries)的传入,将人类不可食用的高原植物转化成能量,也被认为在3.6 ka BP人类全年占据青藏高原的过程中发挥了重要作用[83⇓-85]. ...
... 青藏高原的旧石器时代遗址,除白石崖溶洞遗址(225—45 ka BP、海拔3200 m)[49]外,其他多数为旷野遗址,文化层较薄,揭露的灰坑、火塘等遗迹规模较小,出土遗物数量有限[54,57⇓⇓ -60,89 -90].通常认为,在此阶段人们以在高海拔和低海拔地区长距离移动的方式,间断性访问青藏高原高海拔地区[81,91],动物资源利用方式为纯粹的狩猎经济(图2).从利用的动物种类来看,旧石器中晚期(225—13.1 ka BP)以犀牛(Dicerorhinus)、马科(Equidae)、鹿科(Cervidae)、牛科(Bovidae)等大中型哺乳动物为主,在全新世早中期(10.4—5 ka BP)转变为以羊亚科(Caprinae)/羚羊亚科(Antilopinae)、啮齿类(Rodentia)和狐狸(Vulpes sp.)等中小型动物为主[49,54,56,61]. ...
Agriculture facilitated permanent human occupation of the Tibetan Plateau after 3600 B P
1
2015
... 部分学者认为人群全年占据青藏高原海拔3300 m以上地区的时间不早于7.0 ka BP,且与低海拔农业人群保持密切联系,而人群全年占据海拔4000 m以上地区不早于4.0 ka BP,且与牦牛(Bos grunniens)驯养关系密切[81].来自青藏高原东北部的动植物遗存的证据则表明,3600 a BP麦作农业的传入促进人类大规模占据青藏高原2500 m以上高海拔地区[82].另外,驯化动物,如绵羊(Ovis aries)的传入,将人类不可食用的高原植物转化成能量,也被认为在3.6 ka BP人类全年占据青藏高原的过程中发挥了重要作用[83⇓-85]. ...
The cultural context of biological adaptation to high elevation Tibet
1
2016
... 部分学者认为人群全年占据青藏高原海拔3300 m以上地区的时间不早于7.0 ka BP,且与低海拔农业人群保持密切联系,而人群全年占据海拔4000 m以上地区不早于4.0 ka BP,且与牦牛(Bos grunniens)驯养关系密切[81].来自青藏高原东北部的动植物遗存的证据则表明,3600 a BP麦作农业的传入促进人类大规模占据青藏高原2500 m以上高海拔地区[82].另外,驯化动物,如绵羊(Ovis aries)的传入,将人类不可食用的高原植物转化成能量,也被认为在3.6 ka BP人类全年占据青藏高原的过程中发挥了重要作用[83⇓-85]. ...
The late Upper Paleolithic occupation of the northern Tibetan Plateau margin
1
2006
... 部分学者认为人群全年占据青藏高原海拔3300 m以上地区的时间不早于7.0 ka BP,且与低海拔农业人群保持密切联系,而人群全年占据海拔4000 m以上地区不早于4.0 ka BP,且与牦牛(Bos grunniens)驯养关系密切[81].来自青藏高原东北部的动植物遗存的证据则表明,3600 a BP麦作农业的传入促进人类大规模占据青藏高原2500 m以上高海拔地区[82].另外,驯化动物,如绵羊(Ovis aries)的传入,将人类不可食用的高原植物转化成能量,也被认为在3.6 ka BP人类全年占据青藏高原的过程中发挥了重要作用[83⇓-85]. ...
A short chronology for the peopling of the Tibetan Plateau
1
2007
... 部分学者认为人群全年占据青藏高原海拔3300 m以上地区的时间不早于7.0 ka BP,且与低海拔农业人群保持密切联系,而人群全年占据海拔4000 m以上地区不早于4.0 ka BP,且与牦牛(Bos grunniens)驯养关系密切[81].来自青藏高原东北部的动植物遗存的证据则表明,3600 a BP麦作农业的传入促进人类大规模占据青藏高原2500 m以上高海拔地区[82].另外,驯化动物,如绵羊(Ovis aries)的传入,将人类不可食用的高原植物转化成能量,也被认为在3.6 ka BP人类全年占据青藏高原的过程中发挥了重要作用[83⇓-85]. ...
青海同德县宗日遗址发掘简报
1
1998
... 关于早期人群在青藏高原上开始营定居生活的时间、地点和驱动机制存在不同认识.考古学家根据青海同德宗日遗址和西藏昌都卡若遗址的房址、墓葬、农具、动植物遗存等证据,认为早在5.0 ka BP前后,人类已经定居在海拔超过3000 m的青藏高原东缘地带[68,86];张东菊等认为粟作农业的传播推动史前人类在6.0—4.0 ka BP进入青藏高原东北和东南部的低海拔河谷地带并定居,而后从西方传入的麦作农业促使了人群在3.6 ka BP向更高海拔常年定居[47];同时,根据高原东北部3.6—2.3 ka BP 5个诺木洪文化遗址的动植物考古学证据,学者提出放牧羊和牦牛及大麦种植为主的农牧并重的生产方式,支撑了海拔超过2800 m的诺木洪文化定居的形成[87].最近,陈发虎等将高原定居过程整理出3个阶段,即粟作农业人群至少在5.2 ka BP前进入高原东部低海拔的河谷地带,在4.8 ka BP以后开始定居在高原东部3000 m以上的高海拔区域;3.5 ka BP以来麦作的传入和牧业经济的发展助力人群大规模定居高海拔区域[87].但上述研究,除经过系统发掘的宗日、卡若和小恩达遗址,其他涉及的3000 m以上高海拔考古遗址目前还都缺少一年四季常年定居的直接证据. ...
青海同德县宗日遗址发掘简报
1
1998
... 关于早期人群在青藏高原上开始营定居生活的时间、地点和驱动机制存在不同认识.考古学家根据青海同德宗日遗址和西藏昌都卡若遗址的房址、墓葬、农具、动植物遗存等证据,认为早在5.0 ka BP前后,人类已经定居在海拔超过3000 m的青藏高原东缘地带[68,86];张东菊等认为粟作农业的传播推动史前人类在6.0—4.0 ka BP进入青藏高原东北和东南部的低海拔河谷地带并定居,而后从西方传入的麦作农业促使了人群在3.6 ka BP向更高海拔常年定居[47];同时,根据高原东北部3.6—2.3 ka BP 5个诺木洪文化遗址的动植物考古学证据,学者提出放牧羊和牦牛及大麦种植为主的农牧并重的生产方式,支撑了海拔超过2800 m的诺木洪文化定居的形成[87].最近,陈发虎等将高原定居过程整理出3个阶段,即粟作农业人群至少在5.2 ka BP前进入高原东部低海拔的河谷地带,在4.8 ka BP以后开始定居在高原东部3000 m以上的高海拔区域;3.5 ka BP以来麦作的传入和牧业经济的发展助力人群大规模定居高海拔区域[87].但上述研究,除经过系统发掘的宗日、卡若和小恩达遗址,其他涉及的3000 m以上高海拔考古遗址目前还都缺少一年四季常年定居的直接证据. ...
青藏高原东北部青铜时代中晚期人类对不同海拔环境的适应策略探讨
2
2017
... 关于早期人群在青藏高原上开始营定居生活的时间、地点和驱动机制存在不同认识.考古学家根据青海同德宗日遗址和西藏昌都卡若遗址的房址、墓葬、农具、动植物遗存等证据,认为早在5.0 ka BP前后,人类已经定居在海拔超过3000 m的青藏高原东缘地带[68,86];张东菊等认为粟作农业的传播推动史前人类在6.0—4.0 ka BP进入青藏高原东北和东南部的低海拔河谷地带并定居,而后从西方传入的麦作农业促使了人群在3.6 ka BP向更高海拔常年定居[47];同时,根据高原东北部3.6—2.3 ka BP 5个诺木洪文化遗址的动植物考古学证据,学者提出放牧羊和牦牛及大麦种植为主的农牧并重的生产方式,支撑了海拔超过2800 m的诺木洪文化定居的形成[87].最近,陈发虎等将高原定居过程整理出3个阶段,即粟作农业人群至少在5.2 ka BP前进入高原东部低海拔的河谷地带,在4.8 ka BP以后开始定居在高原东部3000 m以上的高海拔区域;3.5 ka BP以来麦作的传入和牧业经济的发展助力人群大规模定居高海拔区域[87].但上述研究,除经过系统发掘的宗日、卡若和小恩达遗址,其他涉及的3000 m以上高海拔考古遗址目前还都缺少一年四季常年定居的直接证据. ...
... [87].但上述研究,除经过系统发掘的宗日、卡若和小恩达遗址,其他涉及的3000 m以上高海拔考古遗址目前还都缺少一年四季常年定居的直接证据. ...
青藏高原东北部青铜时代中晚期人类对不同海拔环境的适应策略探讨
2
2017
... 关于早期人群在青藏高原上开始营定居生活的时间、地点和驱动机制存在不同认识.考古学家根据青海同德宗日遗址和西藏昌都卡若遗址的房址、墓葬、农具、动植物遗存等证据,认为早在5.0 ka BP前后,人类已经定居在海拔超过3000 m的青藏高原东缘地带[68,86];张东菊等认为粟作农业的传播推动史前人类在6.0—4.0 ka BP进入青藏高原东北和东南部的低海拔河谷地带并定居,而后从西方传入的麦作农业促使了人群在3.6 ka BP向更高海拔常年定居[47];同时,根据高原东北部3.6—2.3 ka BP 5个诺木洪文化遗址的动植物考古学证据,学者提出放牧羊和牦牛及大麦种植为主的农牧并重的生产方式,支撑了海拔超过2800 m的诺木洪文化定居的形成[87].最近,陈发虎等将高原定居过程整理出3个阶段,即粟作农业人群至少在5.2 ka BP前进入高原东部低海拔的河谷地带,在4.8 ka BP以后开始定居在高原东部3000 m以上的高海拔区域;3.5 ka BP以来麦作的传入和牧业经济的发展助力人群大规模定居高海拔区域[87].但上述研究,除经过系统发掘的宗日、卡若和小恩达遗址,其他涉及的3000 m以上高海拔考古遗址目前还都缺少一年四季常年定居的直接证据. ...
... [87].但上述研究,除经过系统发掘的宗日、卡若和小恩达遗址,其他涉及的3000 m以上高海拔考古遗址目前还都缺少一年四季常年定居的直接证据. ...
拓殖高原、永久定居、麦作传入: 对当前青藏高原史前考古重大争论的回应
2
2017
... 以上关于史前人类全年游猎和定居青藏高原的理论大多建立在低地农业人群向高地输出的假设之上,农业的传入自然成为人类可以常年生活在青藏高原高海拔地区的前提.但“人类何时定居高原”和“青藏高原最早的农业”这两个问题是否能等同化[88]?非农业社会是否能够永久定居青藏高原?在青藏高原长久存在的高原狩猎采集者在早期定居和作物传播过程中扮演何种角色[47]?学者们认为,在讨论人类何时定居高原这一问题时,野生动物资源利用的作用被低估,低地河谷农业人群和高原狩猎采集人群之间的关系被忽略[88]. ...
... [88]. ...
拓殖高原、永久定居、麦作传入: 对当前青藏高原史前考古重大争论的回应
2
2017
... 以上关于史前人类全年游猎和定居青藏高原的理论大多建立在低地农业人群向高地输出的假设之上,农业的传入自然成为人类可以常年生活在青藏高原高海拔地区的前提.但“人类何时定居高原”和“青藏高原最早的农业”这两个问题是否能等同化[88]?非农业社会是否能够永久定居青藏高原?在青藏高原长久存在的高原狩猎采集者在早期定居和作物传播过程中扮演何种角色[47]?学者们认为,在讨论人类何时定居高原这一问题时,野生动物资源利用的作用被低估,低地河谷农业人群和高原狩猎采集人群之间的关系被忽略[88]. ...
... [88]. ...
OSL chronology for lacustrine sediments recording high stands of Gahai Lake in Qaidam Basin, northeastern Qinghai-Tibetan Plateau
1
2010
... 青藏高原的旧石器时代遗址,除白石崖溶洞遗址(225—45 ka BP、海拔3200 m)[49]外,其他多数为旷野遗址,文化层较薄,揭露的灰坑、火塘等遗迹规模较小,出土遗物数量有限[54,57⇓⇓ -60,89 -90].通常认为,在此阶段人们以在高海拔和低海拔地区长距离移动的方式,间断性访问青藏高原高海拔地区[81,91],动物资源利用方式为纯粹的狩猎经济(图2).从利用的动物种类来看,旧石器中晚期(225—13.1 ka BP)以犀牛(Dicerorhinus)、马科(Equidae)、鹿科(Cervidae)、牛科(Bovidae)等大中型哺乳动物为主,在全新世早中期(10.4—5 ka BP)转变为以羊亚科(Caprinae)/羚羊亚科(Antilopinae)、啮齿类(Rodentia)和狐狸(Vulpes sp.)等中小型动物为主[49,54,56,61]. ...
Prehistoric human activity and its environmental background in Lake Donggi Cona basin, northeastern Tibetan Plateau
1
2020
... 青藏高原的旧石器时代遗址,除白石崖溶洞遗址(225—45 ka BP、海拔3200 m)[49]外,其他多数为旷野遗址,文化层较薄,揭露的灰坑、火塘等遗迹规模较小,出土遗物数量有限[54,57⇓⇓ -60,89 -90].通常认为,在此阶段人们以在高海拔和低海拔地区长距离移动的方式,间断性访问青藏高原高海拔地区[81,91],动物资源利用方式为纯粹的狩猎经济(图2).从利用的动物种类来看,旧石器中晚期(225—13.1 ka BP)以犀牛(Dicerorhinus)、马科(Equidae)、鹿科(Cervidae)、牛科(Bovidae)等大中型哺乳动物为主,在全新世早中期(10.4—5 ka BP)转变为以羊亚科(Caprinae)/羚羊亚科(Antilopinae)、啮齿类(Rodentia)和狐狸(Vulpes sp.)等中小型动物为主[49,54,56,61]. ...
Winter-to-summer seasonal migration of microlithic human activities on the Qinghai-Tibet Plateau
1
2020
... 青藏高原的旧石器时代遗址,除白石崖溶洞遗址(225—45 ka BP、海拔3200 m)[49]外,其他多数为旷野遗址,文化层较薄,揭露的灰坑、火塘等遗迹规模较小,出土遗物数量有限[54,57⇓⇓ -60,89 -90].通常认为,在此阶段人们以在高海拔和低海拔地区长距离移动的方式,间断性访问青藏高原高海拔地区[81,91],动物资源利用方式为纯粹的狩猎经济(图2).从利用的动物种类来看,旧石器中晚期(225—13.1 ka BP)以犀牛(Dicerorhinus)、马科(Equidae)、鹿科(Cervidae)、牛科(Bovidae)等大中型哺乳动物为主,在全新世早中期(10.4—5 ka BP)转变为以羊亚科(Caprinae)/羚羊亚科(Antilopinae)、啮齿类(Rodentia)和狐狸(Vulpes sp.)等中小型动物为主[49,54,56,61]. ...
Ancient genomes reveal tropical bovid species in the Tibetan Plateau contributed to the prevalence of hunting game until the late Neolithic
1
2020
... 5.5 ka BP前后,青藏高原东北部、川西等高海拔地区开始出现定居遗址,该时期获取动物资源方式呈现出空间上的差异性.一部分遗址出土动物遗存完全为野生动物,如山那树扎(5170—4960 a BP、海拔2312 m)和小恩达遗址[92-93](图2),也有一部分遗址开始出现一定规模的家畜饲养,包括猪(Sus scrofa domesticus)和狗(Canis lupus familiaris),如宗日和卡若遗址[94⇓-96].在横断山区,虽为同一考古学文化,哈休遗址与营盘山遗址出土动物遗存却存在明显差异:海拔较高的哈休遗址(海拔2800 m)出土动物遗存以野生动物为主,家养动物只有狗[97-98];海拔较低的营盘山遗址(海拔1710 m)出土动物遗存则以家养动物为主,主要为家猪及黄牛(Bos taurus),是以定居农业方式为主,畜牧业、渔猎和采集相结合的一种综合经济[97,99],两个遗址表现出动物资源利用在垂向上的差异(图2). ...
The importance of localized hunting of diverse animals to early inhabitants of the Eastern Tibetan Plateau at the Neolithic site of Xiaoenda
1
2019
... 5.5 ka BP前后,青藏高原东北部、川西等高海拔地区开始出现定居遗址,该时期获取动物资源方式呈现出空间上的差异性.一部分遗址出土动物遗存完全为野生动物,如山那树扎(5170—4960 a BP、海拔2312 m)和小恩达遗址[92-93](图2),也有一部分遗址开始出现一定规模的家畜饲养,包括猪(Sus scrofa domesticus)和狗(Canis lupus familiaris),如宗日和卡若遗址[94⇓-96].在横断山区,虽为同一考古学文化,哈休遗址与营盘山遗址出土动物遗存却存在明显差异:海拔较高的哈休遗址(海拔2800 m)出土动物遗存以野生动物为主,家养动物只有狗[97-98];海拔较低的营盘山遗址(海拔1710 m)出土动物遗存则以家养动物为主,主要为家猪及黄牛(Bos taurus),是以定居农业方式为主,畜牧业、渔猎和采集相结合的一种综合经济[97,99],两个遗址表现出动物资源利用在垂向上的差异(图2). ...
宗日文化遗址动物骨骼的研究//河南省文物考古研究所. 动物考古(第1辑)
1
... 5.5 ka BP前后,青藏高原东北部、川西等高海拔地区开始出现定居遗址,该时期获取动物资源方式呈现出空间上的差异性.一部分遗址出土动物遗存完全为野生动物,如山那树扎(5170—4960 a BP、海拔2312 m)和小恩达遗址[92-93](图2),也有一部分遗址开始出现一定规模的家畜饲养,包括猪(Sus scrofa domesticus)和狗(Canis lupus familiaris),如宗日和卡若遗址[94⇓-96].在横断山区,虽为同一考古学文化,哈休遗址与营盘山遗址出土动物遗存却存在明显差异:海拔较高的哈休遗址(海拔2800 m)出土动物遗存以野生动物为主,家养动物只有狗[97-98];海拔较低的营盘山遗址(海拔1710 m)出土动物遗存则以家养动物为主,主要为家猪及黄牛(Bos taurus),是以定居农业方式为主,畜牧业、渔猎和采集相结合的一种综合经济[97,99],两个遗址表现出动物资源利用在垂向上的差异(图2). ...
宗日文化遗址动物骨骼的研究//河南省文物考古研究所. 动物考古(第1辑)
1
... 5.5 ka BP前后,青藏高原东北部、川西等高海拔地区开始出现定居遗址,该时期获取动物资源方式呈现出空间上的差异性.一部分遗址出土动物遗存完全为野生动物,如山那树扎(5170—4960 a BP、海拔2312 m)和小恩达遗址[92-93](图2),也有一部分遗址开始出现一定规模的家畜饲养,包括猪(Sus scrofa domesticus)和狗(Canis lupus familiaris),如宗日和卡若遗址[94⇓-96].在横断山区,虽为同一考古学文化,哈休遗址与营盘山遗址出土动物遗存却存在明显差异:海拔较高的哈休遗址(海拔2800 m)出土动物遗存以野生动物为主,家养动物只有狗[97-98];海拔较低的营盘山遗址(海拔1710 m)出土动物遗存则以家养动物为主,主要为家猪及黄牛(Bos taurus),是以定居农业方式为主,畜牧业、渔猎和采集相结合的一种综合经济[97,99],两个遗址表现出动物资源利用在垂向上的差异(图2). ...
2
2017
... 5.5 ka BP前后,青藏高原东北部、川西等高海拔地区开始出现定居遗址,该时期获取动物资源方式呈现出空间上的差异性.一部分遗址出土动物遗存完全为野生动物,如山那树扎(5170—4960 a BP、海拔2312 m)和小恩达遗址[92-93](图2),也有一部分遗址开始出现一定规模的家畜饲养,包括猪(Sus scrofa domesticus)和狗(Canis lupus familiaris),如宗日和卡若遗址[94⇓-96].在横断山区,虽为同一考古学文化,哈休遗址与营盘山遗址出土动物遗存却存在明显差异:海拔较高的哈休遗址(海拔2800 m)出土动物遗存以野生动物为主,家养动物只有狗[97-98];海拔较低的营盘山遗址(海拔1710 m)出土动物遗存则以家养动物为主,主要为家猪及黄牛(Bos taurus),是以定居农业方式为主,畜牧业、渔猎和采集相结合的一种综合经济[97,99],两个遗址表现出动物资源利用在垂向上的差异(图2). ...
... 在4.5—3.0 ka BP,史前东西方文化交流加强,驯化于西亚的黄牛、山羊(Capra hircus)和绵羊传入至青海省东北部和云南地区,青藏高原地区先民动物资源利用在空间上呈现多元化,牛和羊超过了猪,成为家畜饲养的主流(图2).家羊(绵羊/山羊)在青藏高原最早出土于金蝉口遗址(4.5—3.6 ka BP、海拔2419 m).长宁遗址(4.3—3.6 ka BP、海拔2300 m)出土的动物遗存以家畜黄牛、山羊和绵羊为主,牛、羊所占比均超过了猪,在先民生业经济中占据主要地位[42,95].青藏高原南部拉萨曲贡遗址先民的生业经济以作物种植和放牧牦牛、绵羊的农牧经济为主,以猎取白唇鹿或马鹿、麝、野猪、藏野驴和各种禽类的狩猎经济为辅[70].在高原西部阿里扎达的格布赛鲁遗址早期墓葬(3560—3000 a BP、海拔3800 m)中随葬有山羊、绵羊和黄牛等家养动物,以及盘羊和鹿等野生动物;遗址晚期墓葬(2310—2127 a BP)中出土了山羊、绵羊、黄牛、马、野驴和鱼等.在3.0 ka BP以后,绵羊、山羊、黄牛、马、狗和牦牛等家养动物在青藏高原被广泛畜养,并在人们日常生活和社会仪式中扮演重要角色[100⇓⇓-103](图2). ...
2
2017
... 5.5 ka BP前后,青藏高原东北部、川西等高海拔地区开始出现定居遗址,该时期获取动物资源方式呈现出空间上的差异性.一部分遗址出土动物遗存完全为野生动物,如山那树扎(5170—4960 a BP、海拔2312 m)和小恩达遗址[92-93](图2),也有一部分遗址开始出现一定规模的家畜饲养,包括猪(Sus scrofa domesticus)和狗(Canis lupus familiaris),如宗日和卡若遗址[94⇓-96].在横断山区,虽为同一考古学文化,哈休遗址与营盘山遗址出土动物遗存却存在明显差异:海拔较高的哈休遗址(海拔2800 m)出土动物遗存以野生动物为主,家养动物只有狗[97-98];海拔较低的营盘山遗址(海拔1710 m)出土动物遗存则以家养动物为主,主要为家猪及黄牛(Bos taurus),是以定居农业方式为主,畜牧业、渔猎和采集相结合的一种综合经济[97,99],两个遗址表现出动物资源利用在垂向上的差异(图2). ...
... 在4.5—3.0 ka BP,史前东西方文化交流加强,驯化于西亚的黄牛、山羊(Capra hircus)和绵羊传入至青海省东北部和云南地区,青藏高原地区先民动物资源利用在空间上呈现多元化,牛和羊超过了猪,成为家畜饲养的主流(图2).家羊(绵羊/山羊)在青藏高原最早出土于金蝉口遗址(4.5—3.6 ka BP、海拔2419 m).长宁遗址(4.3—3.6 ka BP、海拔2300 m)出土的动物遗存以家畜黄牛、山羊和绵羊为主,牛、羊所占比均超过了猪,在先民生业经济中占据主要地位[42,95].青藏高原南部拉萨曲贡遗址先民的生业经济以作物种植和放牧牦牛、绵羊的农牧经济为主,以猎取白唇鹿或马鹿、麝、野猪、藏野驴和各种禽类的狩猎经济为辅[70].在高原西部阿里扎达的格布赛鲁遗址早期墓葬(3560—3000 a BP、海拔3800 m)中随葬有山羊、绵羊和黄牛等家养动物,以及盘羊和鹿等野生动物;遗址晚期墓葬(2310—2127 a BP)中出土了山羊、绵羊、黄牛、马、野驴和鱼等.在3.0 ka BP以后,绵羊、山羊、黄牛、马、狗和牦牛等家养动物在青藏高原被广泛畜养,并在人们日常生活和社会仪式中扮演重要角色[100⇓⇓-103](图2). ...
西藏昌都卡若新石器时代遗址动物群
1
1980
... 5.5 ka BP前后,青藏高原东北部、川西等高海拔地区开始出现定居遗址,该时期获取动物资源方式呈现出空间上的差异性.一部分遗址出土动物遗存完全为野生动物,如山那树扎(5170—4960 a BP、海拔2312 m)和小恩达遗址[92-93](图2),也有一部分遗址开始出现一定规模的家畜饲养,包括猪(Sus scrofa domesticus)和狗(Canis lupus familiaris),如宗日和卡若遗址[94⇓-96].在横断山区,虽为同一考古学文化,哈休遗址与营盘山遗址出土动物遗存却存在明显差异:海拔较高的哈休遗址(海拔2800 m)出土动物遗存以野生动物为主,家养动物只有狗[97-98];海拔较低的营盘山遗址(海拔1710 m)出土动物遗存则以家养动物为主,主要为家猪及黄牛(Bos taurus),是以定居农业方式为主,畜牧业、渔猎和采集相结合的一种综合经济[97,99],两个遗址表现出动物资源利用在垂向上的差异(图2). ...
西藏昌都卡若新石器时代遗址动物群
1
1980
... 5.5 ka BP前后,青藏高原东北部、川西等高海拔地区开始出现定居遗址,该时期获取动物资源方式呈现出空间上的差异性.一部分遗址出土动物遗存完全为野生动物,如山那树扎(5170—4960 a BP、海拔2312 m)和小恩达遗址[92-93](图2),也有一部分遗址开始出现一定规模的家畜饲养,包括猪(Sus scrofa domesticus)和狗(Canis lupus familiaris),如宗日和卡若遗址[94⇓-96].在横断山区,虽为同一考古学文化,哈休遗址与营盘山遗址出土动物遗存却存在明显差异:海拔较高的哈休遗址(海拔2800 m)出土动物遗存以野生动物为主,家养动物只有狗[97-98];海拔较低的营盘山遗址(海拔1710 m)出土动物遗存则以家养动物为主,主要为家猪及黄牛(Bos taurus),是以定居农业方式为主,畜牧业、渔猎和采集相结合的一种综合经济[97,99],两个遗址表现出动物资源利用在垂向上的差异(图2). ...
马尔康哈休遗址出土动物骨骼鉴定报告
2
2006
... 5.5 ka BP前后,青藏高原东北部、川西等高海拔地区开始出现定居遗址,该时期获取动物资源方式呈现出空间上的差异性.一部分遗址出土动物遗存完全为野生动物,如山那树扎(5170—4960 a BP、海拔2312 m)和小恩达遗址[92-93](图2),也有一部分遗址开始出现一定规模的家畜饲养,包括猪(Sus scrofa domesticus)和狗(Canis lupus familiaris),如宗日和卡若遗址[94⇓-96].在横断山区,虽为同一考古学文化,哈休遗址与营盘山遗址出土动物遗存却存在明显差异:海拔较高的哈休遗址(海拔2800 m)出土动物遗存以野生动物为主,家养动物只有狗[97-98];海拔较低的营盘山遗址(海拔1710 m)出土动物遗存则以家养动物为主,主要为家猪及黄牛(Bos taurus),是以定居农业方式为主,畜牧业、渔猎和采集相结合的一种综合经济[97,99],两个遗址表现出动物资源利用在垂向上的差异(图2). ...
... [97,99],两个遗址表现出动物资源利用在垂向上的差异(图2). ...
马尔康哈休遗址出土动物骨骼鉴定报告
2
2006
... 5.5 ka BP前后,青藏高原东北部、川西等高海拔地区开始出现定居遗址,该时期获取动物资源方式呈现出空间上的差异性.一部分遗址出土动物遗存完全为野生动物,如山那树扎(5170—4960 a BP、海拔2312 m)和小恩达遗址[92-93](图2),也有一部分遗址开始出现一定规模的家畜饲养,包括猪(Sus scrofa domesticus)和狗(Canis lupus familiaris),如宗日和卡若遗址[94⇓-96].在横断山区,虽为同一考古学文化,哈休遗址与营盘山遗址出土动物遗存却存在明显差异:海拔较高的哈休遗址(海拔2800 m)出土动物遗存以野生动物为主,家养动物只有狗[97-98];海拔较低的营盘山遗址(海拔1710 m)出土动物遗存则以家养动物为主,主要为家猪及黄牛(Bos taurus),是以定居农业方式为主,畜牧业、渔猎和采集相结合的一种综合经济[97,99],两个遗址表现出动物资源利用在垂向上的差异(图2). ...
... [97,99],两个遗址表现出动物资源利用在垂向上的差异(图2). ...
马尔康哈休遗址史前文化与生业: 兼论岷江上游地区马家窑类型的生业方式
1
2015
... 5.5 ka BP前后,青藏高原东北部、川西等高海拔地区开始出现定居遗址,该时期获取动物资源方式呈现出空间上的差异性.一部分遗址出土动物遗存完全为野生动物,如山那树扎(5170—4960 a BP、海拔2312 m)和小恩达遗址[92-93](图2),也有一部分遗址开始出现一定规模的家畜饲养,包括猪(Sus scrofa domesticus)和狗(Canis lupus familiaris),如宗日和卡若遗址[94⇓-96].在横断山区,虽为同一考古学文化,哈休遗址与营盘山遗址出土动物遗存却存在明显差异:海拔较高的哈休遗址(海拔2800 m)出土动物遗存以野生动物为主,家养动物只有狗[97-98];海拔较低的营盘山遗址(海拔1710 m)出土动物遗存则以家养动物为主,主要为家猪及黄牛(Bos taurus),是以定居农业方式为主,畜牧业、渔猎和采集相结合的一种综合经济[97,99],两个遗址表现出动物资源利用在垂向上的差异(图2). ...
马尔康哈休遗址史前文化与生业: 兼论岷江上游地区马家窑类型的生业方式
1
2015
... 5.5 ka BP前后,青藏高原东北部、川西等高海拔地区开始出现定居遗址,该时期获取动物资源方式呈现出空间上的差异性.一部分遗址出土动物遗存完全为野生动物,如山那树扎(5170—4960 a BP、海拔2312 m)和小恩达遗址[92-93](图2),也有一部分遗址开始出现一定规模的家畜饲养,包括猪(Sus scrofa domesticus)和狗(Canis lupus familiaris),如宗日和卡若遗址[94⇓-96].在横断山区,虽为同一考古学文化,哈休遗址与营盘山遗址出土动物遗存却存在明显差异:海拔较高的哈休遗址(海拔2800 m)出土动物遗存以野生动物为主,家养动物只有狗[97-98];海拔较低的营盘山遗址(海拔1710 m)出土动物遗存则以家养动物为主,主要为家猪及黄牛(Bos taurus),是以定居农业方式为主,畜牧业、渔猎和采集相结合的一种综合经济[97,99],两个遗址表现出动物资源利用在垂向上的差异(图2). ...
浅论动物考古学中两种肉量估算方法: 以营盘山遗址出土的动物骨骼为例
1
2009
... 5.5 ka BP前后,青藏高原东北部、川西等高海拔地区开始出现定居遗址,该时期获取动物资源方式呈现出空间上的差异性.一部分遗址出土动物遗存完全为野生动物,如山那树扎(5170—4960 a BP、海拔2312 m)和小恩达遗址[92-93](图2),也有一部分遗址开始出现一定规模的家畜饲养,包括猪(Sus scrofa domesticus)和狗(Canis lupus familiaris),如宗日和卡若遗址[94⇓-96].在横断山区,虽为同一考古学文化,哈休遗址与营盘山遗址出土动物遗存却存在明显差异:海拔较高的哈休遗址(海拔2800 m)出土动物遗存以野生动物为主,家养动物只有狗[97-98];海拔较低的营盘山遗址(海拔1710 m)出土动物遗存则以家养动物为主,主要为家猪及黄牛(Bos taurus),是以定居农业方式为主,畜牧业、渔猎和采集相结合的一种综合经济[97,99],两个遗址表现出动物资源利用在垂向上的差异(图2). ...
浅论动物考古学中两种肉量估算方法: 以营盘山遗址出土的动物骨骼为例
1
2009
... 5.5 ka BP前后,青藏高原东北部、川西等高海拔地区开始出现定居遗址,该时期获取动物资源方式呈现出空间上的差异性.一部分遗址出土动物遗存完全为野生动物,如山那树扎(5170—4960 a BP、海拔2312 m)和小恩达遗址[92-93](图2),也有一部分遗址开始出现一定规模的家畜饲养,包括猪(Sus scrofa domesticus)和狗(Canis lupus familiaris),如宗日和卡若遗址[94⇓-96].在横断山区,虽为同一考古学文化,哈休遗址与营盘山遗址出土动物遗存却存在明显差异:海拔较高的哈休遗址(海拔2800 m)出土动物遗存以野生动物为主,家养动物只有狗[97-98];海拔较低的营盘山遗址(海拔1710 m)出土动物遗存则以家养动物为主,主要为家猪及黄牛(Bos taurus),是以定居农业方式为主,畜牧业、渔猎和采集相结合的一种综合经济[97,99],两个遗址表现出动物资源利用在垂向上的差异(图2). ...
西藏那曲察秀塘祭祀遗址哺乳动物遗存及其意义//河南省文物考古研究所. 动物考古(第1辑)
1
... 在4.5—3.0 ka BP,史前东西方文化交流加强,驯化于西亚的黄牛、山羊(Capra hircus)和绵羊传入至青海省东北部和云南地区,青藏高原地区先民动物资源利用在空间上呈现多元化,牛和羊超过了猪,成为家畜饲养的主流(图2).家羊(绵羊/山羊)在青藏高原最早出土于金蝉口遗址(4.5—3.6 ka BP、海拔2419 m).长宁遗址(4.3—3.6 ka BP、海拔2300 m)出土的动物遗存以家畜黄牛、山羊和绵羊为主,牛、羊所占比均超过了猪,在先民生业经济中占据主要地位[42,95].青藏高原南部拉萨曲贡遗址先民的生业经济以作物种植和放牧牦牛、绵羊的农牧经济为主,以猎取白唇鹿或马鹿、麝、野猪、藏野驴和各种禽类的狩猎经济为辅[70].在高原西部阿里扎达的格布赛鲁遗址早期墓葬(3560—3000 a BP、海拔3800 m)中随葬有山羊、绵羊和黄牛等家养动物,以及盘羊和鹿等野生动物;遗址晚期墓葬(2310—2127 a BP)中出土了山羊、绵羊、黄牛、马、野驴和鱼等.在3.0 ka BP以后,绵羊、山羊、黄牛、马、狗和牦牛等家养动物在青藏高原被广泛畜养,并在人们日常生活和社会仪式中扮演重要角色[100⇓⇓-103](图2). ...
西藏那曲察秀塘祭祀遗址哺乳动物遗存及其意义//河南省文物考古研究所. 动物考古(第1辑)
1
... 在4.5—3.0 ka BP,史前东西方文化交流加强,驯化于西亚的黄牛、山羊(Capra hircus)和绵羊传入至青海省东北部和云南地区,青藏高原地区先民动物资源利用在空间上呈现多元化,牛和羊超过了猪,成为家畜饲养的主流(图2).家羊(绵羊/山羊)在青藏高原最早出土于金蝉口遗址(4.5—3.6 ka BP、海拔2419 m).长宁遗址(4.3—3.6 ka BP、海拔2300 m)出土的动物遗存以家畜黄牛、山羊和绵羊为主,牛、羊所占比均超过了猪,在先民生业经济中占据主要地位[42,95].青藏高原南部拉萨曲贡遗址先民的生业经济以作物种植和放牧牦牛、绵羊的农牧经济为主,以猎取白唇鹿或马鹿、麝、野猪、藏野驴和各种禽类的狩猎经济为辅[70].在高原西部阿里扎达的格布赛鲁遗址早期墓葬(3560—3000 a BP、海拔3800 m)中随葬有山羊、绵羊和黄牛等家养动物,以及盘羊和鹿等野生动物;遗址晚期墓葬(2310—2127 a BP)中出土了山羊、绵羊、黄牛、马、野驴和鱼等.在3.0 ka BP以后,绵羊、山羊、黄牛、马、狗和牦牛等家养动物在青藏高原被广泛畜养,并在人们日常生活和社会仪式中扮演重要角色[100⇓⇓-103](图2). ...
藏北安多布塔雄曲石室墓动物遗存的鉴定分析
1
2015
... 在4.5—3.0 ka BP,史前东西方文化交流加强,驯化于西亚的黄牛、山羊(Capra hircus)和绵羊传入至青海省东北部和云南地区,青藏高原地区先民动物资源利用在空间上呈现多元化,牛和羊超过了猪,成为家畜饲养的主流(图2).家羊(绵羊/山羊)在青藏高原最早出土于金蝉口遗址(4.5—3.6 ka BP、海拔2419 m).长宁遗址(4.3—3.6 ka BP、海拔2300 m)出土的动物遗存以家畜黄牛、山羊和绵羊为主,牛、羊所占比均超过了猪,在先民生业经济中占据主要地位[42,95].青藏高原南部拉萨曲贡遗址先民的生业经济以作物种植和放牧牦牛、绵羊的农牧经济为主,以猎取白唇鹿或马鹿、麝、野猪、藏野驴和各种禽类的狩猎经济为辅[70].在高原西部阿里扎达的格布赛鲁遗址早期墓葬(3560—3000 a BP、海拔3800 m)中随葬有山羊、绵羊和黄牛等家养动物,以及盘羊和鹿等野生动物;遗址晚期墓葬(2310—2127 a BP)中出土了山羊、绵羊、黄牛、马、野驴和鱼等.在3.0 ka BP以后,绵羊、山羊、黄牛、马、狗和牦牛等家养动物在青藏高原被广泛畜养,并在人们日常生活和社会仪式中扮演重要角色[100⇓⇓-103](图2). ...
藏北安多布塔雄曲石室墓动物遗存的鉴定分析
1
2015
... 在4.5—3.0 ka BP,史前东西方文化交流加强,驯化于西亚的黄牛、山羊(Capra hircus)和绵羊传入至青海省东北部和云南地区,青藏高原地区先民动物资源利用在空间上呈现多元化,牛和羊超过了猪,成为家畜饲养的主流(图2).家羊(绵羊/山羊)在青藏高原最早出土于金蝉口遗址(4.5—3.6 ka BP、海拔2419 m).长宁遗址(4.3—3.6 ka BP、海拔2300 m)出土的动物遗存以家畜黄牛、山羊和绵羊为主,牛、羊所占比均超过了猪,在先民生业经济中占据主要地位[42,95].青藏高原南部拉萨曲贡遗址先民的生业经济以作物种植和放牧牦牛、绵羊的农牧经济为主,以猎取白唇鹿或马鹿、麝、野猪、藏野驴和各种禽类的狩猎经济为辅[70].在高原西部阿里扎达的格布赛鲁遗址早期墓葬(3560—3000 a BP、海拔3800 m)中随葬有山羊、绵羊和黄牛等家养动物,以及盘羊和鹿等野生动物;遗址晚期墓葬(2310—2127 a BP)中出土了山羊、绵羊、黄牛、马、野驴和鱼等.在3.0 ka BP以后,绵羊、山羊、黄牛、马、狗和牦牛等家养动物在青藏高原被广泛畜养,并在人们日常生活和社会仪式中扮演重要角色[100⇓⇓-103](图2). ...
西藏西部阿里卡尔东遗址2013年试掘出土动物遗存鉴定与分析
1
2017
... 在4.5—3.0 ka BP,史前东西方文化交流加强,驯化于西亚的黄牛、山羊(Capra hircus)和绵羊传入至青海省东北部和云南地区,青藏高原地区先民动物资源利用在空间上呈现多元化,牛和羊超过了猪,成为家畜饲养的主流(图2).家羊(绵羊/山羊)在青藏高原最早出土于金蝉口遗址(4.5—3.6 ka BP、海拔2419 m).长宁遗址(4.3—3.6 ka BP、海拔2300 m)出土的动物遗存以家畜黄牛、山羊和绵羊为主,牛、羊所占比均超过了猪,在先民生业经济中占据主要地位[42,95].青藏高原南部拉萨曲贡遗址先民的生业经济以作物种植和放牧牦牛、绵羊的农牧经济为主,以猎取白唇鹿或马鹿、麝、野猪、藏野驴和各种禽类的狩猎经济为辅[70].在高原西部阿里扎达的格布赛鲁遗址早期墓葬(3560—3000 a BP、海拔3800 m)中随葬有山羊、绵羊和黄牛等家养动物,以及盘羊和鹿等野生动物;遗址晚期墓葬(2310—2127 a BP)中出土了山羊、绵羊、黄牛、马、野驴和鱼等.在3.0 ka BP以后,绵羊、山羊、黄牛、马、狗和牦牛等家养动物在青藏高原被广泛畜养,并在人们日常生活和社会仪式中扮演重要角色[100⇓⇓-103](图2). ...
西藏西部阿里卡尔东遗址2013年试掘出土动物遗存鉴定与分析
1
2017
... 在4.5—3.0 ka BP,史前东西方文化交流加强,驯化于西亚的黄牛、山羊(Capra hircus)和绵羊传入至青海省东北部和云南地区,青藏高原地区先民动物资源利用在空间上呈现多元化,牛和羊超过了猪,成为家畜饲养的主流(图2).家羊(绵羊/山羊)在青藏高原最早出土于金蝉口遗址(4.5—3.6 ka BP、海拔2419 m).长宁遗址(4.3—3.6 ka BP、海拔2300 m)出土的动物遗存以家畜黄牛、山羊和绵羊为主,牛、羊所占比均超过了猪,在先民生业经济中占据主要地位[42,95].青藏高原南部拉萨曲贡遗址先民的生业经济以作物种植和放牧牦牛、绵羊的农牧经济为主,以猎取白唇鹿或马鹿、麝、野猪、藏野驴和各种禽类的狩猎经济为辅[70].在高原西部阿里扎达的格布赛鲁遗址早期墓葬(3560—3000 a BP、海拔3800 m)中随葬有山羊、绵羊和黄牛等家养动物,以及盘羊和鹿等野生动物;遗址晚期墓葬(2310—2127 a BP)中出土了山羊、绵羊、黄牛、马、野驴和鱼等.在3.0 ka BP以后,绵羊、山羊、黄牛、马、狗和牦牛等家养动物在青藏高原被广泛畜养,并在人们日常生活和社会仪式中扮演重要角色[100⇓⇓-103](图2). ...
Early agropastoral settlement and cultural change in central Tibet in the first millennium BC: Excavations at Bangga
1
2021
... 在4.5—3.0 ka BP,史前东西方文化交流加强,驯化于西亚的黄牛、山羊(Capra hircus)和绵羊传入至青海省东北部和云南地区,青藏高原地区先民动物资源利用在空间上呈现多元化,牛和羊超过了猪,成为家畜饲养的主流(图2).家羊(绵羊/山羊)在青藏高原最早出土于金蝉口遗址(4.5—3.6 ka BP、海拔2419 m).长宁遗址(4.3—3.6 ka BP、海拔2300 m)出土的动物遗存以家畜黄牛、山羊和绵羊为主,牛、羊所占比均超过了猪,在先民生业经济中占据主要地位[42,95].青藏高原南部拉萨曲贡遗址先民的生业经济以作物种植和放牧牦牛、绵羊的农牧经济为主,以猎取白唇鹿或马鹿、麝、野猪、藏野驴和各种禽类的狩猎经济为辅[70].在高原西部阿里扎达的格布赛鲁遗址早期墓葬(3560—3000 a BP、海拔3800 m)中随葬有山羊、绵羊和黄牛等家养动物,以及盘羊和鹿等野生动物;遗址晚期墓葬(2310—2127 a BP)中出土了山羊、绵羊、黄牛、马、野驴和鱼等.在3.0 ka BP以后,绵羊、山羊、黄牛、马、狗和牦牛等家养动物在青藏高原被广泛畜养,并在人们日常生活和社会仪式中扮演重要角色[100⇓⇓-103](图2). ...
The Archaeology of Animals
4
2012
... 考古遗址出土的动物遗存,是理解人类对动物资源利用的主要研究材料.动物考古学研究方法包括种属鉴定、性别和年龄鉴定、死亡年龄结构分析、形态测量、埋藏学分析、定量统计、病理学分析等,在此基础上探讨古代先民对动物资源的开发利用能力,获取、宰杀、运输、加工和消费动物资源的策略和利用模式,复原古代人类的生活环境和资源条件,了解遗址的使用方式(例如季节性使用和延续时间的长短等)、古代堆积的成因、形成过程和受到后堆积过程影响的程度,研究古代家畜的驯化、传播及动物次级产品开发模式和社会发展历程等[104⇓⇓⇓-108]. ...
... 季节性分析是指示遗址是否为全年居住的直接证据,也反映了人类利用自然资源 的策略和能力.温带地区动物的生长、繁殖和行为都受到季节变化的影响,因此遗址出土的动物遗存是进行遗址季节性分析的重要材料.判断方法包括但不限于:① 是否出现只在特定时间出现的物种,包括候鸟、洄游的鱼类、有迁徙习性的哺乳动物和昆虫的蛹等[107,128];② 一些骨骼的形成具有季节性特征,如鹿角生长—脱落的年度循环和雌性鸟类繁殖季节才会形成的髓质骨[104,108,129];③ 动物的出生季节相对固定,通过牙齿和骨骼判断哺乳动物的死亡年龄,可以计算得到死亡季节[108,130];④ 动物的一些部位终生生长并且受到季节变化的影响表现出周期性生长特征,通过观察生长纹可以得知动物的死亡季节,适合研究的材料包括贝壳、鱼类脊椎、鳃盖骨、耳石和哺乳动物牙骨质等[104];⑤通过食草类动物牙齿釉质氧同位素变化判断死亡季节[131].通过以上方法分析每个动物个体的死亡季节,可以得知一个物种的死亡季节分布;通过汇总所有物种所有个体的死亡季节,便可以得到遗址的使用季节.另外由于遗址堆积形成过程中受到多种埋藏学因素的影响,在利用动物遗存讨论获取季节时应尽量排除因自然营力进入遗址堆积的物种,选择明确因人类活动进入遗址堆积的样品,才能对遗址使用的季节性有更为全面、客观的了解[1,43,104]. ...
... [104];⑤通过食草类动物牙齿釉质氧同位素变化判断死亡季节[131].通过以上方法分析每个动物个体的死亡季节,可以得知一个物种的死亡季节分布;通过汇总所有物种所有个体的死亡季节,便可以得到遗址的使用季节.另外由于遗址堆积形成过程中受到多种埋藏学因素的影响,在利用动物遗存讨论获取季节时应尽量排除因自然营力进入遗址堆积的物种,选择明确因人类活动进入遗址堆积的样品,才能对遗址使用的季节性有更为全面、客观的了解[1,43,104]. ...
... ,104]. ...
1
2018
... 考古遗址出土的动物遗存,是理解人类对动物资源利用的主要研究材料.动物考古学研究方法包括种属鉴定、性别和年龄鉴定、死亡年龄结构分析、形态测量、埋藏学分析、定量统计、病理学分析等,在此基础上探讨古代先民对动物资源的开发利用能力,获取、宰杀、运输、加工和消费动物资源的策略和利用模式,复原古代人类的生活环境和资源条件,了解遗址的使用方式(例如季节性使用和延续时间的长短等)、古代堆积的成因、形成过程和受到后堆积过程影响的程度,研究古代家畜的驯化、传播及动物次级产品开发模式和社会发展历程等[104⇓⇓⇓-108]. ...
1
1994
... 考古遗址出土的动物遗存,是理解人类对动物资源利用的主要研究材料.动物考古学研究方法包括种属鉴定、性别和年龄鉴定、死亡年龄结构分析、形态测量、埋藏学分析、定量统计、病理学分析等,在此基础上探讨古代先民对动物资源的开发利用能力,获取、宰杀、运输、加工和消费动物资源的策略和利用模式,复原古代人类的生活环境和资源条件,了解遗址的使用方式(例如季节性使用和延续时间的长短等)、古代堆积的成因、形成过程和受到后堆积过程影响的程度,研究古代家畜的驯化、传播及动物次级产品开发模式和社会发展历程等[104⇓⇓⇓-108]. ...
2
2008
... 考古遗址出土的动物遗存,是理解人类对动物资源利用的主要研究材料.动物考古学研究方法包括种属鉴定、性别和年龄鉴定、死亡年龄结构分析、形态测量、埋藏学分析、定量统计、病理学分析等,在此基础上探讨古代先民对动物资源的开发利用能力,获取、宰杀、运输、加工和消费动物资源的策略和利用模式,复原古代人类的生活环境和资源条件,了解遗址的使用方式(例如季节性使用和延续时间的长短等)、古代堆积的成因、形成过程和受到后堆积过程影响的程度,研究古代家畜的驯化、传播及动物次级产品开发模式和社会发展历程等[104⇓⇓⇓-108]. ...
... 季节性分析是指示遗址是否为全年居住的直接证据,也反映了人类利用自然资源 的策略和能力.温带地区动物的生长、繁殖和行为都受到季节变化的影响,因此遗址出土的动物遗存是进行遗址季节性分析的重要材料.判断方法包括但不限于:① 是否出现只在特定时间出现的物种,包括候鸟、洄游的鱼类、有迁徙习性的哺乳动物和昆虫的蛹等[107,128];② 一些骨骼的形成具有季节性特征,如鹿角生长—脱落的年度循环和雌性鸟类繁殖季节才会形成的髓质骨[104,108,129];③ 动物的出生季节相对固定,通过牙齿和骨骼判断哺乳动物的死亡年龄,可以计算得到死亡季节[108,130];④ 动物的一些部位终生生长并且受到季节变化的影响表现出周期性生长特征,通过观察生长纹可以得知动物的死亡季节,适合研究的材料包括贝壳、鱼类脊椎、鳃盖骨、耳石和哺乳动物牙骨质等[104];⑤通过食草类动物牙齿釉质氧同位素变化判断死亡季节[131].通过以上方法分析每个动物个体的死亡季节,可以得知一个物种的死亡季节分布;通过汇总所有物种所有个体的死亡季节,便可以得到遗址的使用季节.另外由于遗址堆积形成过程中受到多种埋藏学因素的影响,在利用动物遗存讨论获取季节时应尽量排除因自然营力进入遗址堆积的物种,选择明确因人类活动进入遗址堆积的样品,才能对遗址使用的季节性有更为全面、客观的了解[1,43,104]. ...
3
2008
... 考古遗址出土的动物遗存,是理解人类对动物资源利用的主要研究材料.动物考古学研究方法包括种属鉴定、性别和年龄鉴定、死亡年龄结构分析、形态测量、埋藏学分析、定量统计、病理学分析等,在此基础上探讨古代先民对动物资源的开发利用能力,获取、宰杀、运输、加工和消费动物资源的策略和利用模式,复原古代人类的生活环境和资源条件,了解遗址的使用方式(例如季节性使用和延续时间的长短等)、古代堆积的成因、形成过程和受到后堆积过程影响的程度,研究古代家畜的驯化、传播及动物次级产品开发模式和社会发展历程等[104⇓⇓⇓-108]. ...
... 季节性分析是指示遗址是否为全年居住的直接证据,也反映了人类利用自然资源 的策略和能力.温带地区动物的生长、繁殖和行为都受到季节变化的影响,因此遗址出土的动物遗存是进行遗址季节性分析的重要材料.判断方法包括但不限于:① 是否出现只在特定时间出现的物种,包括候鸟、洄游的鱼类、有迁徙习性的哺乳动物和昆虫的蛹等[107,128];② 一些骨骼的形成具有季节性特征,如鹿角生长—脱落的年度循环和雌性鸟类繁殖季节才会形成的髓质骨[104,108,129];③ 动物的出生季节相对固定,通过牙齿和骨骼判断哺乳动物的死亡年龄,可以计算得到死亡季节[108,130];④ 动物的一些部位终生生长并且受到季节变化的影响表现出周期性生长特征,通过观察生长纹可以得知动物的死亡季节,适合研究的材料包括贝壳、鱼类脊椎、鳃盖骨、耳石和哺乳动物牙骨质等[104];⑤通过食草类动物牙齿釉质氧同位素变化判断死亡季节[131].通过以上方法分析每个动物个体的死亡季节,可以得知一个物种的死亡季节分布;通过汇总所有物种所有个体的死亡季节,便可以得到遗址的使用季节.另外由于遗址堆积形成过程中受到多种埋藏学因素的影响,在利用动物遗存讨论获取季节时应尽量排除因自然营力进入遗址堆积的物种,选择明确因人类活动进入遗址堆积的样品,才能对遗址使用的季节性有更为全面、客观的了解[1,43,104]. ...
... [108,130];④ 动物的一些部位终生生长并且受到季节变化的影响表现出周期性生长特征,通过观察生长纹可以得知动物的死亡季节,适合研究的材料包括贝壳、鱼类脊椎、鳃盖骨、耳石和哺乳动物牙骨质等[104];⑤通过食草类动物牙齿釉质氧同位素变化判断死亡季节[131].通过以上方法分析每个动物个体的死亡季节,可以得知一个物种的死亡季节分布;通过汇总所有物种所有个体的死亡季节,便可以得到遗址的使用季节.另外由于遗址堆积形成过程中受到多种埋藏学因素的影响,在利用动物遗存讨论获取季节时应尽量排除因自然营力进入遗址堆积的物种,选择明确因人类活动进入遗址堆积的样品,才能对遗址使用的季节性有更为全面、客观的了解[1,43,104]. ...
3
1992
... 对遗址出土的动物遗存进行物种鉴定是动物考古研究的基础,需要大量现生动物骨骼样品作为比对标本.目前已知的青藏高原现生动物共1100余种,其中哺乳类190余种,鸟类817种,爬行类55种,两栖类45种,鱼类159种[109⇓⇓⇓-113].青藏高原动物区系具有生物多样性丰富、高原特有种比例高的特点,如在羌塘高原和可可西里栖息的29种哺乳动物中,高原特有种有11种,包括多种世界上独有的有蹄动物,如野牦牛(Bos mutus)、藏羚羊(Pantholops hodgsonii)和藏野驴(Equus kiang)[113].目前青藏高原动物的标本主要存放于博物馆等研究机构,受限于库房场地及动物学研究问题等因素,往往仅保存有头骨而缺少颅后骨,并不适用于考古遗存的研究;已有的骨骼图谱也缺少对高原动物如野牦牛、白唇鹿(Przewalskium albirostris)和藏野驴的描述,动物考古学中形态学的研究也刚刚起步[114].因此,建立和完善青藏高原现生动物骨骼形态数据库至关重要. ...
... 其次,BBM技术鉴定结果的准确性依赖于参考数据库的质量.构建完善的物种DNA条形码的标准数据库、物种信息库、信息共享和应用平台有利于BBM鉴定准确度的提高[127].据统计,青藏高原记录的现生鱼类、鸟类和哺乳动物分别为159种、817种和190余种[109⇓⇓-112].以NCBI(National Center for Biotechnology Information、 https://www.ncbi.nlm.nih.gov/)数据共享平台为参考,本文对青藏高原的149种哺乳动物、152种鱼和234种鸟(主要为涉禽和游禽)的线粒体DNA(mtDNA)数据库构建情况进行了统计(表2).结果表明,其中119种哺乳动物、112种鱼和152种鸟类具有完整的mtDNA参考序列,分别占各类动物统计物种数的78.87%、73.68%和64.96%;无mtDNA数据的动物主要为蝙蝠科(Vespertilionidae)、鼠科(Muridae)、鸠鸽科(Columbidae)、夜鹰科(Caprimulgidae)等物种,这些物种在青藏高原考古遗址中通常也较为少见,因此本文认为,目前青藏高原动物mtDNA数据库平台基本可以支撑BBM数据的分析,但后续仍需进行补充和完善. ...
... 对于各遗址,需要根据遗址出土动物遗存特异性选取相应的研究方法.青藏高原 是亚洲多条大江大河的发源地,且湖泊广布,面积大于10 km²的湖泊共计417个,总面积约44993.4 km² [132],鱼类资源丰富[109].已有考古遗址出土的野生哺乳动物也较为丰富(图2),因此以下两种方法可作为该区域遗址季节性判断的主要参考途径. ...
3
1992
... 对遗址出土的动物遗存进行物种鉴定是动物考古研究的基础,需要大量现生动物骨骼样品作为比对标本.目前已知的青藏高原现生动物共1100余种,其中哺乳类190余种,鸟类817种,爬行类55种,两栖类45种,鱼类159种[109⇓⇓⇓-113].青藏高原动物区系具有生物多样性丰富、高原特有种比例高的特点,如在羌塘高原和可可西里栖息的29种哺乳动物中,高原特有种有11种,包括多种世界上独有的有蹄动物,如野牦牛(Bos mutus)、藏羚羊(Pantholops hodgsonii)和藏野驴(Equus kiang)[113].目前青藏高原动物的标本主要存放于博物馆等研究机构,受限于库房场地及动物学研究问题等因素,往往仅保存有头骨而缺少颅后骨,并不适用于考古遗存的研究;已有的骨骼图谱也缺少对高原动物如野牦牛、白唇鹿(Przewalskium albirostris)和藏野驴的描述,动物考古学中形态学的研究也刚刚起步[114].因此,建立和完善青藏高原现生动物骨骼形态数据库至关重要. ...
... 其次,BBM技术鉴定结果的准确性依赖于参考数据库的质量.构建完善的物种DNA条形码的标准数据库、物种信息库、信息共享和应用平台有利于BBM鉴定准确度的提高[127].据统计,青藏高原记录的现生鱼类、鸟类和哺乳动物分别为159种、817种和190余种[109⇓⇓-112].以NCBI(National Center for Biotechnology Information、 https://www.ncbi.nlm.nih.gov/)数据共享平台为参考,本文对青藏高原的149种哺乳动物、152种鱼和234种鸟(主要为涉禽和游禽)的线粒体DNA(mtDNA)数据库构建情况进行了统计(表2).结果表明,其中119种哺乳动物、112种鱼和152种鸟类具有完整的mtDNA参考序列,分别占各类动物统计物种数的78.87%、73.68%和64.96%;无mtDNA数据的动物主要为蝙蝠科(Vespertilionidae)、鼠科(Muridae)、鸠鸽科(Columbidae)、夜鹰科(Caprimulgidae)等物种,这些物种在青藏高原考古遗址中通常也较为少见,因此本文认为,目前青藏高原动物mtDNA数据库平台基本可以支撑BBM数据的分析,但后续仍需进行补充和完善. ...
... 对于各遗址,需要根据遗址出土动物遗存特异性选取相应的研究方法.青藏高原 是亚洲多条大江大河的发源地,且湖泊广布,面积大于10 km²的湖泊共计417个,总面积约44993.4 km² [132],鱼类资源丰富[109].已有考古遗址出土的野生哺乳动物也较为丰富(图2),因此以下两种方法可作为该区域遗址季节性判断的主要参考途径. ...
青藏高原生物多样性保护研究
2
2004
... 对遗址出土的动物遗存进行物种鉴定是动物考古研究的基础,需要大量现生动物骨骼样品作为比对标本.目前已知的青藏高原现生动物共1100余种,其中哺乳类190余种,鸟类817种,爬行类55种,两栖类45种,鱼类159种[109⇓⇓⇓-113].青藏高原动物区系具有生物多样性丰富、高原特有种比例高的特点,如在羌塘高原和可可西里栖息的29种哺乳动物中,高原特有种有11种,包括多种世界上独有的有蹄动物,如野牦牛(Bos mutus)、藏羚羊(Pantholops hodgsonii)和藏野驴(Equus kiang)[113].目前青藏高原动物的标本主要存放于博物馆等研究机构,受限于库房场地及动物学研究问题等因素,往往仅保存有头骨而缺少颅后骨,并不适用于考古遗存的研究;已有的骨骼图谱也缺少对高原动物如野牦牛、白唇鹿(Przewalskium albirostris)和藏野驴的描述,动物考古学中形态学的研究也刚刚起步[114].因此,建立和完善青藏高原现生动物骨骼形态数据库至关重要. ...
... 其次,BBM技术鉴定结果的准确性依赖于参考数据库的质量.构建完善的物种DNA条形码的标准数据库、物种信息库、信息共享和应用平台有利于BBM鉴定准确度的提高[127].据统计,青藏高原记录的现生鱼类、鸟类和哺乳动物分别为159种、817种和190余种[109⇓⇓-112].以NCBI(National Center for Biotechnology Information、 https://www.ncbi.nlm.nih.gov/)数据共享平台为参考,本文对青藏高原的149种哺乳动物、152种鱼和234种鸟(主要为涉禽和游禽)的线粒体DNA(mtDNA)数据库构建情况进行了统计(表2).结果表明,其中119种哺乳动物、112种鱼和152种鸟类具有完整的mtDNA参考序列,分别占各类动物统计物种数的78.87%、73.68%和64.96%;无mtDNA数据的动物主要为蝙蝠科(Vespertilionidae)、鼠科(Muridae)、鸠鸽科(Columbidae)、夜鹰科(Caprimulgidae)等物种,这些物种在青藏高原考古遗址中通常也较为少见,因此本文认为,目前青藏高原动物mtDNA数据库平台基本可以支撑BBM数据的分析,但后续仍需进行补充和完善. ...
青藏高原生物多样性保护研究
2
2004
... 对遗址出土的动物遗存进行物种鉴定是动物考古研究的基础,需要大量现生动物骨骼样品作为比对标本.目前已知的青藏高原现生动物共1100余种,其中哺乳类190余种,鸟类817种,爬行类55种,两栖类45种,鱼类159种[109⇓⇓⇓-113].青藏高原动物区系具有生物多样性丰富、高原特有种比例高的特点,如在羌塘高原和可可西里栖息的29种哺乳动物中,高原特有种有11种,包括多种世界上独有的有蹄动物,如野牦牛(Bos mutus)、藏羚羊(Pantholops hodgsonii)和藏野驴(Equus kiang)[113].目前青藏高原动物的标本主要存放于博物馆等研究机构,受限于库房场地及动物学研究问题等因素,往往仅保存有头骨而缺少颅后骨,并不适用于考古遗存的研究;已有的骨骼图谱也缺少对高原动物如野牦牛、白唇鹿(Przewalskium albirostris)和藏野驴的描述,动物考古学中形态学的研究也刚刚起步[114].因此,建立和完善青藏高原现生动物骨骼形态数据库至关重要. ...
... 其次,BBM技术鉴定结果的准确性依赖于参考数据库的质量.构建完善的物种DNA条形码的标准数据库、物种信息库、信息共享和应用平台有利于BBM鉴定准确度的提高[127].据统计,青藏高原记录的现生鱼类、鸟类和哺乳动物分别为159种、817种和190余种[109⇓⇓-112].以NCBI(National Center for Biotechnology Information、 https://www.ncbi.nlm.nih.gov/)数据共享平台为参考,本文对青藏高原的149种哺乳动物、152种鱼和234种鸟(主要为涉禽和游禽)的线粒体DNA(mtDNA)数据库构建情况进行了统计(表2).结果表明,其中119种哺乳动物、112种鱼和152种鸟类具有完整的mtDNA参考序列,分别占各类动物统计物种数的78.87%、73.68%和64.96%;无mtDNA数据的动物主要为蝙蝠科(Vespertilionidae)、鼠科(Muridae)、鸠鸽科(Columbidae)、夜鹰科(Caprimulgidae)等物种,这些物种在青藏高原考古遗址中通常也较为少见,因此本文认为,目前青藏高原动物mtDNA数据库平台基本可以支撑BBM数据的分析,但后续仍需进行补充和完善. ...
2
2013
... 对遗址出土的动物遗存进行物种鉴定是动物考古研究的基础,需要大量现生动物骨骼样品作为比对标本.目前已知的青藏高原现生动物共1100余种,其中哺乳类190余种,鸟类817种,爬行类55种,两栖类45种,鱼类159种[109⇓⇓⇓-113].青藏高原动物区系具有生物多样性丰富、高原特有种比例高的特点,如在羌塘高原和可可西里栖息的29种哺乳动物中,高原特有种有11种,包括多种世界上独有的有蹄动物,如野牦牛(Bos mutus)、藏羚羊(Pantholops hodgsonii)和藏野驴(Equus kiang)[113].目前青藏高原动物的标本主要存放于博物馆等研究机构,受限于库房场地及动物学研究问题等因素,往往仅保存有头骨而缺少颅后骨,并不适用于考古遗存的研究;已有的骨骼图谱也缺少对高原动物如野牦牛、白唇鹿(Przewalskium albirostris)和藏野驴的描述,动物考古学中形态学的研究也刚刚起步[114].因此,建立和完善青藏高原现生动物骨骼形态数据库至关重要. ...
... 其次,BBM技术鉴定结果的准确性依赖于参考数据库的质量.构建完善的物种DNA条形码的标准数据库、物种信息库、信息共享和应用平台有利于BBM鉴定准确度的提高[127].据统计,青藏高原记录的现生鱼类、鸟类和哺乳动物分别为159种、817种和190余种[109⇓⇓-112].以NCBI(National Center for Biotechnology Information、 https://www.ncbi.nlm.nih.gov/)数据共享平台为参考,本文对青藏高原的149种哺乳动物、152种鱼和234种鸟(主要为涉禽和游禽)的线粒体DNA(mtDNA)数据库构建情况进行了统计(表2).结果表明,其中119种哺乳动物、112种鱼和152种鸟类具有完整的mtDNA参考序列,分别占各类动物统计物种数的78.87%、73.68%和64.96%;无mtDNA数据的动物主要为蝙蝠科(Vespertilionidae)、鼠科(Muridae)、鸠鸽科(Columbidae)、夜鹰科(Caprimulgidae)等物种,这些物种在青藏高原考古遗址中通常也较为少见,因此本文认为,目前青藏高原动物mtDNA数据库平台基本可以支撑BBM数据的分析,但后续仍需进行补充和完善. ...
2
2013
... 对遗址出土的动物遗存进行物种鉴定是动物考古研究的基础,需要大量现生动物骨骼样品作为比对标本.目前已知的青藏高原现生动物共1100余种,其中哺乳类190余种,鸟类817种,爬行类55种,两栖类45种,鱼类159种[109⇓⇓⇓-113].青藏高原动物区系具有生物多样性丰富、高原特有种比例高的特点,如在羌塘高原和可可西里栖息的29种哺乳动物中,高原特有种有11种,包括多种世界上独有的有蹄动物,如野牦牛(Bos mutus)、藏羚羊(Pantholops hodgsonii)和藏野驴(Equus kiang)[113].目前青藏高原动物的标本主要存放于博物馆等研究机构,受限于库房场地及动物学研究问题等因素,往往仅保存有头骨而缺少颅后骨,并不适用于考古遗存的研究;已有的骨骼图谱也缺少对高原动物如野牦牛、白唇鹿(Przewalskium albirostris)和藏野驴的描述,动物考古学中形态学的研究也刚刚起步[114].因此,建立和完善青藏高原现生动物骨骼形态数据库至关重要. ...
... 其次,BBM技术鉴定结果的准确性依赖于参考数据库的质量.构建完善的物种DNA条形码的标准数据库、物种信息库、信息共享和应用平台有利于BBM鉴定准确度的提高[127].据统计,青藏高原记录的现生鱼类、鸟类和哺乳动物分别为159种、817种和190余种[109⇓⇓-112].以NCBI(National Center for Biotechnology Information、 https://www.ncbi.nlm.nih.gov/)数据共享平台为参考,本文对青藏高原的149种哺乳动物、152种鱼和234种鸟(主要为涉禽和游禽)的线粒体DNA(mtDNA)数据库构建情况进行了统计(表2).结果表明,其中119种哺乳动物、112种鱼和152种鸟类具有完整的mtDNA参考序列,分别占各类动物统计物种数的78.87%、73.68%和64.96%;无mtDNA数据的动物主要为蝙蝠科(Vespertilionidae)、鼠科(Muridae)、鸠鸽科(Columbidae)、夜鹰科(Caprimulgidae)等物种,这些物种在青藏高原考古遗址中通常也较为少见,因此本文认为,目前青藏高原动物mtDNA数据库平台基本可以支撑BBM数据的分析,但后续仍需进行补充和完善. ...
2
2005
... 对遗址出土的动物遗存进行物种鉴定是动物考古研究的基础,需要大量现生动物骨骼样品作为比对标本.目前已知的青藏高原现生动物共1100余种,其中哺乳类190余种,鸟类817种,爬行类55种,两栖类45种,鱼类159种[109⇓⇓⇓-113].青藏高原动物区系具有生物多样性丰富、高原特有种比例高的特点,如在羌塘高原和可可西里栖息的29种哺乳动物中,高原特有种有11种,包括多种世界上独有的有蹄动物,如野牦牛(Bos mutus)、藏羚羊(Pantholops hodgsonii)和藏野驴(Equus kiang)[113].目前青藏高原动物的标本主要存放于博物馆等研究机构,受限于库房场地及动物学研究问题等因素,往往仅保存有头骨而缺少颅后骨,并不适用于考古遗存的研究;已有的骨骼图谱也缺少对高原动物如野牦牛、白唇鹿(Przewalskium albirostris)和藏野驴的描述,动物考古学中形态学的研究也刚刚起步[114].因此,建立和完善青藏高原现生动物骨骼形态数据库至关重要. ...
... 其次,BBM技术鉴定结果的准确性依赖于参考数据库的质量.构建完善的物种DNA条形码的标准数据库、物种信息库、信息共享和应用平台有利于BBM鉴定准确度的提高[127].据统计,青藏高原记录的现生鱼类、鸟类和哺乳动物分别为159种、817种和190余种[109⇓⇓-112].以NCBI(National Center for Biotechnology Information、 https://www.ncbi.nlm.nih.gov/)数据共享平台为参考,本文对青藏高原的149种哺乳动物、152种鱼和234种鸟(主要为涉禽和游禽)的线粒体DNA(mtDNA)数据库构建情况进行了统计(表2).结果表明,其中119种哺乳动物、112种鱼和152种鸟类具有完整的mtDNA参考序列,分别占各类动物统计物种数的78.87%、73.68%和64.96%;无mtDNA数据的动物主要为蝙蝠科(Vespertilionidae)、鼠科(Muridae)、鸠鸽科(Columbidae)、夜鹰科(Caprimulgidae)等物种,这些物种在青藏高原考古遗址中通常也较为少见,因此本文认为,目前青藏高原动物mtDNA数据库平台基本可以支撑BBM数据的分析,但后续仍需进行补充和完善. ...
2
2005
... 对遗址出土的动物遗存进行物种鉴定是动物考古研究的基础,需要大量现生动物骨骼样品作为比对标本.目前已知的青藏高原现生动物共1100余种,其中哺乳类190余种,鸟类817种,爬行类55种,两栖类45种,鱼类159种[109⇓⇓⇓-113].青藏高原动物区系具有生物多样性丰富、高原特有种比例高的特点,如在羌塘高原和可可西里栖息的29种哺乳动物中,高原特有种有11种,包括多种世界上独有的有蹄动物,如野牦牛(Bos mutus)、藏羚羊(Pantholops hodgsonii)和藏野驴(Equus kiang)[113].目前青藏高原动物的标本主要存放于博物馆等研究机构,受限于库房场地及动物学研究问题等因素,往往仅保存有头骨而缺少颅后骨,并不适用于考古遗存的研究;已有的骨骼图谱也缺少对高原动物如野牦牛、白唇鹿(Przewalskium albirostris)和藏野驴的描述,动物考古学中形态学的研究也刚刚起步[114].因此,建立和完善青藏高原现生动物骨骼形态数据库至关重要. ...
... 其次,BBM技术鉴定结果的准确性依赖于参考数据库的质量.构建完善的物种DNA条形码的标准数据库、物种信息库、信息共享和应用平台有利于BBM鉴定准确度的提高[127].据统计,青藏高原记录的现生鱼类、鸟类和哺乳动物分别为159种、817种和190余种[109⇓⇓-112].以NCBI(National Center for Biotechnology Information、 https://www.ncbi.nlm.nih.gov/)数据共享平台为参考,本文对青藏高原的149种哺乳动物、152种鱼和234种鸟(主要为涉禽和游禽)的线粒体DNA(mtDNA)数据库构建情况进行了统计(表2).结果表明,其中119种哺乳动物、112种鱼和152种鸟类具有完整的mtDNA参考序列,分别占各类动物统计物种数的78.87%、73.68%和64.96%;无mtDNA数据的动物主要为蝙蝠科(Vespertilionidae)、鼠科(Muridae)、鸠鸽科(Columbidae)、夜鹰科(Caprimulgidae)等物种,这些物种在青藏高原考古遗址中通常也较为少见,因此本文认为,目前青藏高原动物mtDNA数据库平台基本可以支撑BBM数据的分析,但后续仍需进行补充和完善. ...
2
2018
... 对遗址出土的动物遗存进行物种鉴定是动物考古研究的基础,需要大量现生动物骨骼样品作为比对标本.目前已知的青藏高原现生动物共1100余种,其中哺乳类190余种,鸟类817种,爬行类55种,两栖类45种,鱼类159种[109⇓⇓⇓-113].青藏高原动物区系具有生物多样性丰富、高原特有种比例高的特点,如在羌塘高原和可可西里栖息的29种哺乳动物中,高原特有种有11种,包括多种世界上独有的有蹄动物,如野牦牛(Bos mutus)、藏羚羊(Pantholops hodgsonii)和藏野驴(Equus kiang)[113].目前青藏高原动物的标本主要存放于博物馆等研究机构,受限于库房场地及动物学研究问题等因素,往往仅保存有头骨而缺少颅后骨,并不适用于考古遗存的研究;已有的骨骼图谱也缺少对高原动物如野牦牛、白唇鹿(Przewalskium albirostris)和藏野驴的描述,动物考古学中形态学的研究也刚刚起步[114].因此,建立和完善青藏高原现生动物骨骼形态数据库至关重要. ...
... [113].目前青藏高原动物的标本主要存放于博物馆等研究机构,受限于库房场地及动物学研究问题等因素,往往仅保存有头骨而缺少颅后骨,并不适用于考古遗存的研究;已有的骨骼图谱也缺少对高原动物如野牦牛、白唇鹿(Przewalskium albirostris)和藏野驴的描述,动物考古学中形态学的研究也刚刚起步[114].因此,建立和完善青藏高原现生动物骨骼形态数据库至关重要. ...
2
2018
... 对遗址出土的动物遗存进行物种鉴定是动物考古研究的基础,需要大量现生动物骨骼样品作为比对标本.目前已知的青藏高原现生动物共1100余种,其中哺乳类190余种,鸟类817种,爬行类55种,两栖类45种,鱼类159种[109⇓⇓⇓-113].青藏高原动物区系具有生物多样性丰富、高原特有种比例高的特点,如在羌塘高原和可可西里栖息的29种哺乳动物中,高原特有种有11种,包括多种世界上独有的有蹄动物,如野牦牛(Bos mutus)、藏羚羊(Pantholops hodgsonii)和藏野驴(Equus kiang)[113].目前青藏高原动物的标本主要存放于博物馆等研究机构,受限于库房场地及动物学研究问题等因素,往往仅保存有头骨而缺少颅后骨,并不适用于考古遗存的研究;已有的骨骼图谱也缺少对高原动物如野牦牛、白唇鹿(Przewalskium albirostris)和藏野驴的描述,动物考古学中形态学的研究也刚刚起步[114].因此,建立和完善青藏高原现生动物骨骼形态数据库至关重要. ...
... [113].目前青藏高原动物的标本主要存放于博物馆等研究机构,受限于库房场地及动物学研究问题等因素,往往仅保存有头骨而缺少颅后骨,并不适用于考古遗存的研究;已有的骨骼图谱也缺少对高原动物如野牦牛、白唇鹿(Przewalskium albirostris)和藏野驴的描述,动物考古学中形态学的研究也刚刚起步[114].因此,建立和完善青藏高原现生动物骨骼形态数据库至关重要. ...
Morphological and metric criteria for identifying postcranial skeletal remains of modern and archaeological Caprinae and Antilopinae in the northeast Tibetan Plateau and adjacent areas
1
2020
... 对遗址出土的动物遗存进行物种鉴定是动物考古研究的基础,需要大量现生动物骨骼样品作为比对标本.目前已知的青藏高原现生动物共1100余种,其中哺乳类190余种,鸟类817种,爬行类55种,两栖类45种,鱼类159种[109⇓⇓⇓-113].青藏高原动物区系具有生物多样性丰富、高原特有种比例高的特点,如在羌塘高原和可可西里栖息的29种哺乳动物中,高原特有种有11种,包括多种世界上独有的有蹄动物,如野牦牛(Bos mutus)、藏羚羊(Pantholops hodgsonii)和藏野驴(Equus kiang)[113].目前青藏高原动物的标本主要存放于博物馆等研究机构,受限于库房场地及动物学研究问题等因素,往往仅保存有头骨而缺少颅后骨,并不适用于考古遗存的研究;已有的骨骼图谱也缺少对高原动物如野牦牛、白唇鹿(Przewalskium albirostris)和藏野驴的描述,动物考古学中形态学的研究也刚刚起步[114].因此,建立和完善青藏高原现生动物骨骼形态数据库至关重要. ...
Scrapheap challenge: A novel bulk-bone metabarcoding method to investigate ancient DNA in faunal assemblages
2
2013
... 大批量骨骼宏条形码技术(Bulk Bone Metabarcoding, BBM)是指通过宏条形码分析方法,一次性检测混合骨骼样本中所包含的物种分类信息的技术[115].BBM整合了DNA条形码和高通量测序技术,通过提取混合骨骼样品中的DNA,使用特异性PCR(Polymerase Chain Reaction)引物进行扩增,并对扩增产物进行测序分析,来实现混合骨骼样品的物种鉴定,以分析考古遗址的动物群组合(图3)[116]. ...
... 2013年BBM首次被应用到澳大利亚西南部Devil's Lair and Tunnel洞穴遗址出土的骨骼碎片粉末样品(Bulk-bone Powder Samples)分析中,得到了其中哺乳动物、鸟类和爬行动物的物种构成[115].而后,BBM方法在马达加斯加、澳大利亚和新西兰等热带地区,以及小高加索山脉和巴西地区被应用,对时间跨度在42000—100 a BP的骨骼样品进行了分析,对古代人群利用动物资源的种类和生业经济策略、生物多样性变化、环境变化和人类活动对生态环境的影响等问题进行了讨论[117⇓⇓⇓-121].BBM所需样品量少,可以确定形态上不可鉴定的动物碎骨,鉴定出的物种往往包含化石记录中最丰富的物种,且可以鉴定出一系列灭绝的物种,因此,BBM被认为是一种创新、高效地识别考古遗址动物群的方法,具有准确性和灵敏性的特点,可以广泛应用[122]. ...
Towards next‐generation biodiversity assessment using DNA metabarcoding
4
2012
... 大批量骨骼宏条形码技术(Bulk Bone Metabarcoding, BBM)是指通过宏条形码分析方法,一次性检测混合骨骼样本中所包含的物种分类信息的技术[115].BBM整合了DNA条形码和高通量测序技术,通过提取混合骨骼样品中的DNA,使用特异性PCR(Polymerase Chain Reaction)引物进行扩增,并对扩增产物进行测序分析,来实现混合骨骼样品的物种鉴定,以分析考古遗址的动物群组合(图3)[116]. ...
... 首先,Metabarcoding方法依赖于PCR扩增,因而找到适合于扩增特定分类群的、短的、可变的DNA区域的条形码对整个研究的成功起到至关重要的作用的.扩增的基因片段在不同物种间至少要有几个百分点的碱基对差异,在同一物种间该片段的差异则应当很小[116].如目前多使用16S rRNA基因条形码引物用于哺乳动物,12S rRNA引物用于鸟类和鱼类进行特异性扩增(表1)[122-123]. ...
... 最后,相比于现代样品的宏条形码研究,BBM常常用于分析古代样品,而样品中的古DNA高度降解碎片化严重,长度的限制也降低了BBM技术对物种分类的分辨率.BBM只能对生物群体如鱼类、鸟类和哺乳动物中的单独一类进行扩增,所以难以在一个DNA提取物中评估多个类群物种的相对比例[116];并且,从单个细胞器标记识别物种也存在一些问题,如对于进化过程中相对年轻的物种,其在标记的条形码上特异性不高,或者物种间基因渐渗的存在,都可能导致物种的错误识别[116]. ...
... [116]. ...
Multi-analytical approach to zooarchaeological assemblages elucidates Late Holocene coastal lifeways in southwest Madagascar
1
2018
... 2013年BBM首次被应用到澳大利亚西南部Devil's Lair and Tunnel洞穴遗址出土的骨骼碎片粉末样品(Bulk-bone Powder Samples)分析中,得到了其中哺乳动物、鸟类和爬行动物的物种构成[115].而后,BBM方法在马达加斯加、澳大利亚和新西兰等热带地区,以及小高加索山脉和巴西地区被应用,对时间跨度在42000—100 a BP的骨骼样品进行了分析,对古代人群利用动物资源的种类和生业经济策略、生物多样性变化、环境变化和人类活动对生态环境的影响等问题进行了讨论[117⇓⇓⇓-121].BBM所需样品量少,可以确定形态上不可鉴定的动物碎骨,鉴定出的物种往往包含化石记录中最丰富的物种,且可以鉴定出一系列灭绝的物种,因此,BBM被认为是一种创新、高效地识别考古遗址动物群的方法,具有准确性和灵敏性的特点,可以广泛应用[122]. ...
Subsistence practices, past biodiversity, and anthropogenic impacts revealed by New Zealand-wide ancient DNA survey
1
2018
... 2013年BBM首次被应用到澳大利亚西南部Devil's Lair and Tunnel洞穴遗址出土的骨骼碎片粉末样品(Bulk-bone Powder Samples)分析中,得到了其中哺乳动物、鸟类和爬行动物的物种构成[115].而后,BBM方法在马达加斯加、澳大利亚和新西兰等热带地区,以及小高加索山脉和巴西地区被应用,对时间跨度在42000—100 a BP的骨骼样品进行了分析,对古代人群利用动物资源的种类和生业经济策略、生物多样性变化、环境变化和人类活动对生态环境的影响等问题进行了讨论[117⇓⇓⇓-121].BBM所需样品量少,可以确定形态上不可鉴定的动物碎骨,鉴定出的物种往往包含化石记录中最丰富的物种,且可以鉴定出一系列灭绝的物种,因此,BBM被认为是一种创新、高效地识别考古遗址动物群的方法,具有准确性和灵敏性的特点,可以广泛应用[122]. ...
Ancient DNA from bulk bone reveals past genetic diversity of vertebrate fauna on Kangaroo Island, Australia
1
2021
... 2013年BBM首次被应用到澳大利亚西南部Devil's Lair and Tunnel洞穴遗址出土的骨骼碎片粉末样品(Bulk-bone Powder Samples)分析中,得到了其中哺乳动物、鸟类和爬行动物的物种构成[115].而后,BBM方法在马达加斯加、澳大利亚和新西兰等热带地区,以及小高加索山脉和巴西地区被应用,对时间跨度在42000—100 a BP的骨骼样品进行了分析,对古代人群利用动物资源的种类和生业经济策略、生物多样性变化、环境变化和人类活动对生态环境的影响等问题进行了讨论[117⇓⇓⇓-121].BBM所需样品量少,可以确定形态上不可鉴定的动物碎骨,鉴定出的物种往往包含化石记录中最丰富的物种,且可以鉴定出一系列灭绝的物种,因此,BBM被认为是一种创新、高效地识别考古遗址动物群的方法,具有准确性和灵敏性的特点,可以广泛应用[122]. ...
Ancient DNA preserved in small bone fragments from the PW Lund collection
1
2021
... 2013年BBM首次被应用到澳大利亚西南部Devil's Lair and Tunnel洞穴遗址出土的骨骼碎片粉末样品(Bulk-bone Powder Samples)分析中,得到了其中哺乳动物、鸟类和爬行动物的物种构成[115].而后,BBM方法在马达加斯加、澳大利亚和新西兰等热带地区,以及小高加索山脉和巴西地区被应用,对时间跨度在42000—100 a BP的骨骼样品进行了分析,对古代人群利用动物资源的种类和生业经济策略、生物多样性变化、环境变化和人类活动对生态环境的影响等问题进行了讨论[117⇓⇓⇓-121].BBM所需样品量少,可以确定形态上不可鉴定的动物碎骨,鉴定出的物种往往包含化石记录中最丰富的物种,且可以鉴定出一系列灭绝的物种,因此,BBM被认为是一种创新、高效地识别考古遗址动物群的方法,具有准确性和灵敏性的特点,可以广泛应用[122]. ...
Ancient DNA shows high faunal diversity in the Lesser Caucasus during the Late Pleistocene
1
2019
... 2013年BBM首次被应用到澳大利亚西南部Devil's Lair and Tunnel洞穴遗址出土的骨骼碎片粉末样品(Bulk-bone Powder Samples)分析中,得到了其中哺乳动物、鸟类和爬行动物的物种构成[115].而后,BBM方法在马达加斯加、澳大利亚和新西兰等热带地区,以及小高加索山脉和巴西地区被应用,对时间跨度在42000—100 a BP的骨骼样品进行了分析,对古代人群利用动物资源的种类和生业经济策略、生物多样性变化、环境变化和人类活动对生态环境的影响等问题进行了讨论[117⇓⇓⇓-121].BBM所需样品量少,可以确定形态上不可鉴定的动物碎骨,鉴定出的物种往往包含化石记录中最丰富的物种,且可以鉴定出一系列灭绝的物种,因此,BBM被认为是一种创新、高效地识别考古遗址动物群的方法,具有准确性和灵敏性的特点,可以广泛应用[122]. ...
A critical evaluation of how ancient DNA bulk bone metabarcoding complements traditional morphological analysis of fossil assemblages
2
2015
... 2013年BBM首次被应用到澳大利亚西南部Devil's Lair and Tunnel洞穴遗址出土的骨骼碎片粉末样品(Bulk-bone Powder Samples)分析中,得到了其中哺乳动物、鸟类和爬行动物的物种构成[115].而后,BBM方法在马达加斯加、澳大利亚和新西兰等热带地区,以及小高加索山脉和巴西地区被应用,对时间跨度在42000—100 a BP的骨骼样品进行了分析,对古代人群利用动物资源的种类和生业经济策略、生物多样性变化、环境变化和人类活动对生态环境的影响等问题进行了讨论[117⇓⇓⇓-121].BBM所需样品量少,可以确定形态上不可鉴定的动物碎骨,鉴定出的物种往往包含化石记录中最丰富的物种,且可以鉴定出一系列灭绝的物种,因此,BBM被认为是一种创新、高效地识别考古遗址动物群的方法,具有准确性和灵敏性的特点,可以广泛应用[122]. ...
... 首先,Metabarcoding方法依赖于PCR扩增,因而找到适合于扩增特定分类群的、短的、可变的DNA区域的条形码对整个研究的成功起到至关重要的作用的.扩增的基因片段在不同物种间至少要有几个百分点的碱基对差异,在同一物种间该片段的差异则应当很小[116].如目前多使用16S rRNA基因条形码引物用于哺乳动物,12S rRNA引物用于鸟类和鱼类进行特异性扩增(表1)[122-123]. ...
An assessment of ancient DNA preservation in Holocene-Pleistocene fossil bone excavated from the world heritage Naracoorte Caves, South Australia
1
2016
... 首先,Metabarcoding方法依赖于PCR扩增,因而找到适合于扩增特定分类群的、短的、可变的DNA区域的条形码对整个研究的成功起到至关重要的作用的.扩增的基因片段在不同物种间至少要有几个百分点的碱基对差异,在同一物种间该片段的差异则应当很小[116].如目前多使用16S rRNA基因条形码引物用于哺乳动物,12S rRNA引物用于鸟类和鱼类进行特异性扩增(表1)[122-123]. ...
Ancient DNA sequences reveal unsuspected phylogenetic relationships within New Zealand wrens (Acanthisittidae)
2
1994
... Primers for specific amplification in bulk-bone metabarcoding
Tab. 1 | 名称 | | 序列(5'-3') | 目标
分类群 | 基因片段 | 参考文献 | 扩增长度(bp) | 退火温度(°C) |
| 12SAH | 正向 | CTGGGATTAGATACCCCACTAT | 鸟类 | 12S rRNA | [124] | 229~235 | 57 |
| 逆向 | CCTTGACCTGTCTTGTTAGC | 鸟类 | 12S rRNA | [124] | 229~235 | 57 |
| Mam16S | 正向 | CGGTTGGGGTGACCTCGGA | 哺乳类 | 16S rRNA | [125] | 89~115 | 57 |
| 逆向 | GCTGTTATCCCTAGGGTAACT | 哺乳类 | 16S rRNA | [125] | 89~115 | 57 |
| 12S-V5 | 正向 | ACTGGGATTAGATACCCC | 鱼类 | 12S rRNA | [126] | 106 | 58 |
| 逆向 | TAGAACAGGCTCCTCTAG | 鱼类 | 12S rRNA | [126] | 106 | 58 |
其次,BBM技术鉴定结果的准确性依赖于参考数据库的质量.构建完善的物种DNA条形码的标准数据库、物种信息库、信息共享和应用平台有利于BBM鉴定准确度的提高[127].据统计,青藏高原记录的现生鱼类、鸟类和哺乳动物分别为159种、817种和190余种[109⇓⇓-112].以NCBI(National Center for Biotechnology Information、 https://www.ncbi.nlm.nih.gov/)数据共享平台为参考,本文对青藏高原的149种哺乳动物、152种鱼和234种鸟(主要为涉禽和游禽)的线粒体DNA(mtDNA)数据库构建情况进行了统计(表2).结果表明,其中119种哺乳动物、112种鱼和152种鸟类具有完整的mtDNA参考序列,分别占各类动物统计物种数的78.87%、73.68%和64.96%;无mtDNA数据的动物主要为蝙蝠科(Vespertilionidae)、鼠科(Muridae)、鸠鸽科(Columbidae)、夜鹰科(Caprimulgidae)等物种,这些物种在青藏高原考古遗址中通常也较为少见,因此本文认为,目前青藏高原动物mtDNA数据库平台基本可以支撑BBM数据的分析,但后续仍需进行补充和完善. ...
... [
124]
229~235 | 57 | | Mam16S | 正向 | CGGTTGGGGTGACCTCGGA | 哺乳类 | 16S rRNA | [125] | 89~115 | 57 |
| 逆向 | GCTGTTATCCCTAGGGTAACT | 哺乳类 | 16S rRNA | [125] | 89~115 | 57 |
| 12S-V5 | 正向 | ACTGGGATTAGATACCCC | 鱼类 | 12S rRNA | [126] | 106 | 58 |
| 逆向 | TAGAACAGGCTCCTCTAG | 鱼类 | 12S rRNA | [126] | 106 | 58 |
其次,BBM技术鉴定结果的准确性依赖于参考数据库的质量.构建完善的物种DNA条形码的标准数据库、物种信息库、信息共享和应用平台有利于BBM鉴定准确度的提高[127].据统计,青藏高原记录的现生鱼类、鸟类和哺乳动物分别为159种、817种和190余种[109⇓⇓-112].以NCBI(National Center for Biotechnology Information、 https://www.ncbi.nlm.nih.gov/)数据共享平台为参考,本文对青藏高原的149种哺乳动物、152种鱼和234种鸟(主要为涉禽和游禽)的线粒体DNA(mtDNA)数据库构建情况进行了统计(表2).结果表明,其中119种哺乳动物、112种鱼和152种鸟类具有完整的mtDNA参考序列,分别占各类动物统计物种数的78.87%、73.68%和64.96%;无mtDNA数据的动物主要为蝙蝠科(Vespertilionidae)、鼠科(Muridae)、鸠鸽科(Columbidae)、夜鹰科(Caprimulgidae)等物种,这些物种在青藏高原考古遗址中通常也较为少见,因此本文认为,目前青藏高原动物mtDNA数据库平台基本可以支撑BBM数据的分析,但后续仍需进行补充和完善. ...
Reproducibility of ancient DNA sequences from extinct Pleistocene fauna
2
1996
... Primers for specific amplification in bulk-bone metabarcoding
Tab. 1 | 名称 | | 序列(5'-3') | 目标
分类群 | 基因片段 | 参考文献 | 扩增长度(bp) | 退火温度(°C) |
| 12SAH | 正向 | CTGGGATTAGATACCCCACTAT | 鸟类 | 12S rRNA | [124] | 229~235 | 57 |
| 逆向 | CCTTGACCTGTCTTGTTAGC | 鸟类 | 12S rRNA | [124] | 229~235 | 57 |
| Mam16S | 正向 | CGGTTGGGGTGACCTCGGA | 哺乳类 | 16S rRNA | [125] | 89~115 | 57 |
| 逆向 | GCTGTTATCCCTAGGGTAACT | 哺乳类 | 16S rRNA | [125] | 89~115 | 57 |
| 12S-V5 | 正向 | ACTGGGATTAGATACCCC | 鱼类 | 12S rRNA | [126] | 106 | 58 |
| 逆向 | TAGAACAGGCTCCTCTAG | 鱼类 | 12S rRNA | [126] | 106 | 58 |
其次,BBM技术鉴定结果的准确性依赖于参考数据库的质量.构建完善的物种DNA条形码的标准数据库、物种信息库、信息共享和应用平台有利于BBM鉴定准确度的提高[127].据统计,青藏高原记录的现生鱼类、鸟类和哺乳动物分别为159种、817种和190余种[109⇓⇓-112].以NCBI(National Center for Biotechnology Information、 https://www.ncbi.nlm.nih.gov/)数据共享平台为参考,本文对青藏高原的149种哺乳动物、152种鱼和234种鸟(主要为涉禽和游禽)的线粒体DNA(mtDNA)数据库构建情况进行了统计(表2).结果表明,其中119种哺乳动物、112种鱼和152种鸟类具有完整的mtDNA参考序列,分别占各类动物统计物种数的78.87%、73.68%和64.96%;无mtDNA数据的动物主要为蝙蝠科(Vespertilionidae)、鼠科(Muridae)、鸠鸽科(Columbidae)、夜鹰科(Caprimulgidae)等物种,这些物种在青藏高原考古遗址中通常也较为少见,因此本文认为,目前青藏高原动物mtDNA数据库平台基本可以支撑BBM数据的分析,但后续仍需进行补充和完善. ...
... [
125]
89~115 | 57 | | 12S-V5 | 正向 | ACTGGGATTAGATACCCC | 鱼类 | 12S rRNA | [126] | 106 | 58 |
| 逆向 | TAGAACAGGCTCCTCTAG | 鱼类 | 12S rRNA | [126] | 106 | 58 |
其次,BBM技术鉴定结果的准确性依赖于参考数据库的质量.构建完善的物种DNA条形码的标准数据库、物种信息库、信息共享和应用平台有利于BBM鉴定准确度的提高[127].据统计,青藏高原记录的现生鱼类、鸟类和哺乳动物分别为159种、817种和190余种[109⇓⇓-112].以NCBI(National Center for Biotechnology Information、 https://www.ncbi.nlm.nih.gov/)数据共享平台为参考,本文对青藏高原的149种哺乳动物、152种鱼和234种鸟(主要为涉禽和游禽)的线粒体DNA(mtDNA)数据库构建情况进行了统计(表2).结果表明,其中119种哺乳动物、112种鱼和152种鸟类具有完整的mtDNA参考序列,分别占各类动物统计物种数的78.87%、73.68%和64.96%;无mtDNA数据的动物主要为蝙蝠科(Vespertilionidae)、鼠科(Muridae)、鸠鸽科(Columbidae)、夜鹰科(Caprimulgidae)等物种,这些物种在青藏高原考古遗址中通常也较为少见,因此本文认为,目前青藏高原动物mtDNA数据库平台基本可以支撑BBM数据的分析,但后续仍需进行补充和完善. ...
EcoPrimers: Inference of new DNA barcode markers from whole genome sequence analysis
2
2011
... Primers for specific amplification in bulk-bone metabarcoding
Tab. 1 | 名称 | | 序列(5'-3') | 目标
分类群 | 基因片段 | 参考文献 | 扩增长度(bp) | 退火温度(°C) |
| 12SAH | 正向 | CTGGGATTAGATACCCCACTAT | 鸟类 | 12S rRNA | [124] | 229~235 | 57 |
| 逆向 | CCTTGACCTGTCTTGTTAGC | 鸟类 | 12S rRNA | [124] | 229~235 | 57 |
| Mam16S | 正向 | CGGTTGGGGTGACCTCGGA | 哺乳类 | 16S rRNA | [125] | 89~115 | 57 |
| 逆向 | GCTGTTATCCCTAGGGTAACT | 哺乳类 | 16S rRNA | [125] | 89~115 | 57 |
| 12S-V5 | 正向 | ACTGGGATTAGATACCCC | 鱼类 | 12S rRNA | [126] | 106 | 58 |
| 逆向 | TAGAACAGGCTCCTCTAG | 鱼类 | 12S rRNA | [126] | 106 | 58 |
其次,BBM技术鉴定结果的准确性依赖于参考数据库的质量.构建完善的物种DNA条形码的标准数据库、物种信息库、信息共享和应用平台有利于BBM鉴定准确度的提高[127].据统计,青藏高原记录的现生鱼类、鸟类和哺乳动物分别为159种、817种和190余种[109⇓⇓-112].以NCBI(National Center for Biotechnology Information、 https://www.ncbi.nlm.nih.gov/)数据共享平台为参考,本文对青藏高原的149种哺乳动物、152种鱼和234种鸟(主要为涉禽和游禽)的线粒体DNA(mtDNA)数据库构建情况进行了统计(表2).结果表明,其中119种哺乳动物、112种鱼和152种鸟类具有完整的mtDNA参考序列,分别占各类动物统计物种数的78.87%、73.68%和64.96%;无mtDNA数据的动物主要为蝙蝠科(Vespertilionidae)、鼠科(Muridae)、鸠鸽科(Columbidae)、夜鹰科(Caprimulgidae)等物种,这些物种在青藏高原考古遗址中通常也较为少见,因此本文认为,目前青藏高原动物mtDNA数据库平台基本可以支撑BBM数据的分析,但后续仍需进行补充和完善. ...
... [
126]
106 | 58 | 其次,BBM技术鉴定结果的准确性依赖于参考数据库的质量.构建完善的物种DNA条形码的标准数据库、物种信息库、信息共享和应用平台有利于BBM鉴定准确度的提高[127].据统计,青藏高原记录的现生鱼类、鸟类和哺乳动物分别为159种、817种和190余种[109⇓⇓-112].以NCBI(National Center for Biotechnology Information、 https://www.ncbi.nlm.nih.gov/)数据共享平台为参考,本文对青藏高原的149种哺乳动物、152种鱼和234种鸟(主要为涉禽和游禽)的线粒体DNA(mtDNA)数据库构建情况进行了统计(表2).结果表明,其中119种哺乳动物、112种鱼和152种鸟类具有完整的mtDNA参考序列,分别占各类动物统计物种数的78.87%、73.68%和64.96%;无mtDNA数据的动物主要为蝙蝠科(Vespertilionidae)、鼠科(Muridae)、鸠鸽科(Columbidae)、夜鹰科(Caprimulgidae)等物种,这些物种在青藏高原考古遗址中通常也较为少见,因此本文认为,目前青藏高原动物mtDNA数据库平台基本可以支撑BBM数据的分析,但后续仍需进行补充和完善. ...
环境DNA metabarcoding及其在生态学研究中的应用
1
2016
... 其次,BBM技术鉴定结果的准确性依赖于参考数据库的质量.构建完善的物种DNA条形码的标准数据库、物种信息库、信息共享和应用平台有利于BBM鉴定准确度的提高[127].据统计,青藏高原记录的现生鱼类、鸟类和哺乳动物分别为159种、817种和190余种[109⇓⇓-112].以NCBI(National Center for Biotechnology Information、 https://www.ncbi.nlm.nih.gov/)数据共享平台为参考,本文对青藏高原的149种哺乳动物、152种鱼和234种鸟(主要为涉禽和游禽)的线粒体DNA(mtDNA)数据库构建情况进行了统计(表2).结果表明,其中119种哺乳动物、112种鱼和152种鸟类具有完整的mtDNA参考序列,分别占各类动物统计物种数的78.87%、73.68%和64.96%;无mtDNA数据的动物主要为蝙蝠科(Vespertilionidae)、鼠科(Muridae)、鸠鸽科(Columbidae)、夜鹰科(Caprimulgidae)等物种,这些物种在青藏高原考古遗址中通常也较为少见,因此本文认为,目前青藏高原动物mtDNA数据库平台基本可以支撑BBM数据的分析,但后续仍需进行补充和完善. ...
环境DNA metabarcoding及其在生态学研究中的应用
1
2016
... 其次,BBM技术鉴定结果的准确性依赖于参考数据库的质量.构建完善的物种DNA条形码的标准数据库、物种信息库、信息共享和应用平台有利于BBM鉴定准确度的提高[127].据统计,青藏高原记录的现生鱼类、鸟类和哺乳动物分别为159种、817种和190余种[109⇓⇓-112].以NCBI(National Center for Biotechnology Information、 https://www.ncbi.nlm.nih.gov/)数据共享平台为参考,本文对青藏高原的149种哺乳动物、152种鱼和234种鸟(主要为涉禽和游禽)的线粒体DNA(mtDNA)数据库构建情况进行了统计(表2).结果表明,其中119种哺乳动物、112种鱼和152种鸟类具有完整的mtDNA参考序列,分别占各类动物统计物种数的78.87%、73.68%和64.96%;无mtDNA数据的动物主要为蝙蝠科(Vespertilionidae)、鼠科(Muridae)、鸠鸽科(Columbidae)、夜鹰科(Caprimulgidae)等物种,这些物种在青藏高原考古遗址中通常也较为少见,因此本文认为,目前青藏高原动物mtDNA数据库平台基本可以支撑BBM数据的分析,但后续仍需进行补充和完善. ...
Preservation of seasonally abundant waterfowl? Analysis of faunal remains from middens at the Pre-Pottery Neolithic a site of Shubayqa 6 in northeast Jordan
1
2020
... 季节性分析是指示遗址是否为全年居住的直接证据,也反映了人类利用自然资源 的策略和能力.温带地区动物的生长、繁殖和行为都受到季节变化的影响,因此遗址出土的动物遗存是进行遗址季节性分析的重要材料.判断方法包括但不限于:① 是否出现只在特定时间出现的物种,包括候鸟、洄游的鱼类、有迁徙习性的哺乳动物和昆虫的蛹等[107,128];② 一些骨骼的形成具有季节性特征,如鹿角生长—脱落的年度循环和雌性鸟类繁殖季节才会形成的髓质骨[104,108,129];③ 动物的出生季节相对固定,通过牙齿和骨骼判断哺乳动物的死亡年龄,可以计算得到死亡季节[108,130];④ 动物的一些部位终生生长并且受到季节变化的影响表现出周期性生长特征,通过观察生长纹可以得知动物的死亡季节,适合研究的材料包括贝壳、鱼类脊椎、鳃盖骨、耳石和哺乳动物牙骨质等[104];⑤通过食草类动物牙齿釉质氧同位素变化判断死亡季节[131].通过以上方法分析每个动物个体的死亡季节,可以得知一个物种的死亡季节分布;通过汇总所有物种所有个体的死亡季节,便可以得到遗址的使用季节.另外由于遗址堆积形成过程中受到多种埋藏学因素的影响,在利用动物遗存讨论获取季节时应尽量排除因自然营力进入遗址堆积的物种,选择明确因人类活动进入遗址堆积的样品,才能对遗址使用的季节性有更为全面、客观的了解[1,43,104]. ...
1
2009
... 季节性分析是指示遗址是否为全年居住的直接证据,也反映了人类利用自然资源 的策略和能力.温带地区动物的生长、繁殖和行为都受到季节变化的影响,因此遗址出土的动物遗存是进行遗址季节性分析的重要材料.判断方法包括但不限于:① 是否出现只在特定时间出现的物种,包括候鸟、洄游的鱼类、有迁徙习性的哺乳动物和昆虫的蛹等[107,128];② 一些骨骼的形成具有季节性特征,如鹿角生长—脱落的年度循环和雌性鸟类繁殖季节才会形成的髓质骨[104,108,129];③ 动物的出生季节相对固定,通过牙齿和骨骼判断哺乳动物的死亡年龄,可以计算得到死亡季节[108,130];④ 动物的一些部位终生生长并且受到季节变化的影响表现出周期性生长特征,通过观察生长纹可以得知动物的死亡季节,适合研究的材料包括贝壳、鱼类脊椎、鳃盖骨、耳石和哺乳动物牙骨质等[104];⑤通过食草类动物牙齿釉质氧同位素变化判断死亡季节[131].通过以上方法分析每个动物个体的死亡季节,可以得知一个物种的死亡季节分布;通过汇总所有物种所有个体的死亡季节,便可以得到遗址的使用季节.另外由于遗址堆积形成过程中受到多种埋藏学因素的影响,在利用动物遗存讨论获取季节时应尽量排除因自然营力进入遗址堆积的物种,选择明确因人类活动进入遗址堆积的样品,才能对遗址使用的季节性有更为全面、客观的了解[1,43,104]. ...
2
2005
... 季节性分析是指示遗址是否为全年居住的直接证据,也反映了人类利用自然资源 的策略和能力.温带地区动物的生长、繁殖和行为都受到季节变化的影响,因此遗址出土的动物遗存是进行遗址季节性分析的重要材料.判断方法包括但不限于:① 是否出现只在特定时间出现的物种,包括候鸟、洄游的鱼类、有迁徙习性的哺乳动物和昆虫的蛹等[107,128];② 一些骨骼的形成具有季节性特征,如鹿角生长—脱落的年度循环和雌性鸟类繁殖季节才会形成的髓质骨[104,108,129];③ 动物的出生季节相对固定,通过牙齿和骨骼判断哺乳动物的死亡年龄,可以计算得到死亡季节[108,130];④ 动物的一些部位终生生长并且受到季节变化的影响表现出周期性生长特征,通过观察生长纹可以得知动物的死亡季节,适合研究的材料包括贝壳、鱼类脊椎、鳃盖骨、耳石和哺乳动物牙骨质等[104];⑤通过食草类动物牙齿釉质氧同位素变化判断死亡季节[131].通过以上方法分析每个动物个体的死亡季节,可以得知一个物种的死亡季节分布;通过汇总所有物种所有个体的死亡季节,便可以得到遗址的使用季节.另外由于遗址堆积形成过程中受到多种埋藏学因素的影响,在利用动物遗存讨论获取季节时应尽量排除因自然营力进入遗址堆积的物种,选择明确因人类活动进入遗址堆积的样品,才能对遗址使用的季节性有更为全面、客观的了解[1,43,104]. ...
... 动物考古学判断哺乳动物死亡年龄的方法,主要包括骨干骺端愈合情况、牙齿萌出次序和磨蚀程度、齿冠高度测量等[130].牙釉质坚硬易保存,因此经常通过牙齿萌出顺序和磨蚀程度来判断动物的死亡年龄,结合该物种的出生时间可以间接推断动物的死亡季节.但是由于分析得到的死亡年龄和动物的出生季节往往是分布区间,这种方法在判断动物死亡季节时精度较低[133]. ...
The combined use of oxygen isotopes and microwear in sheep teeth to elucidate seasonal management of domestic herds: The case study of ?atalh?yük, central Anatolia
1
2012
... 季节性分析是指示遗址是否为全年居住的直接证据,也反映了人类利用自然资源 的策略和能力.温带地区动物的生长、繁殖和行为都受到季节变化的影响,因此遗址出土的动物遗存是进行遗址季节性分析的重要材料.判断方法包括但不限于:① 是否出现只在特定时间出现的物种,包括候鸟、洄游的鱼类、有迁徙习性的哺乳动物和昆虫的蛹等[107,128];② 一些骨骼的形成具有季节性特征,如鹿角生长—脱落的年度循环和雌性鸟类繁殖季节才会形成的髓质骨[104,108,129];③ 动物的出生季节相对固定,通过牙齿和骨骼判断哺乳动物的死亡年龄,可以计算得到死亡季节[108,130];④ 动物的一些部位终生生长并且受到季节变化的影响表现出周期性生长特征,通过观察生长纹可以得知动物的死亡季节,适合研究的材料包括贝壳、鱼类脊椎、鳃盖骨、耳石和哺乳动物牙骨质等[104];⑤通过食草类动物牙齿釉质氧同位素变化判断死亡季节[131].通过以上方法分析每个动物个体的死亡季节,可以得知一个物种的死亡季节分布;通过汇总所有物种所有个体的死亡季节,便可以得到遗址的使用季节.另外由于遗址堆积形成过程中受到多种埋藏学因素的影响,在利用动物遗存讨论获取季节时应尽量排除因自然营力进入遗址堆积的物种,选择明确因人类活动进入遗址堆积的样品,才能对遗址使用的季节性有更为全面、客观的了解[1,43,104]. ...
近40年青藏高原湖泊面积变化遥感分析
1
2014
... 对于各遗址,需要根据遗址出土动物遗存特异性选取相应的研究方法.青藏高原 是亚洲多条大江大河的发源地,且湖泊广布,面积大于10 km²的湖泊共计417个,总面积约44993.4 km² [132],鱼类资源丰富[109].已有考古遗址出土的野生哺乳动物也较为丰富(图2),因此以下两种方法可作为该区域遗址季节性判断的主要参考途径. ...
近40年青藏高原湖泊面积变化遥感分析
1
2014
... 对于各遗址,需要根据遗址出土动物遗存特异性选取相应的研究方法.青藏高原 是亚洲多条大江大河的发源地,且湖泊广布,面积大于10 km²的湖泊共计417个,总面积约44993.4 km² [132],鱼类资源丰富[109].已有考古遗址出土的野生哺乳动物也较为丰富(图2),因此以下两种方法可作为该区域遗址季节性判断的主要参考途径. ...
垩质年轮法在旧石器时代动物考古学中的应用
1
2014
... 动物考古学判断哺乳动物死亡年龄的方法,主要包括骨干骺端愈合情况、牙齿萌出次序和磨蚀程度、齿冠高度测量等[130].牙釉质坚硬易保存,因此经常通过牙齿萌出顺序和磨蚀程度来判断动物的死亡年龄,结合该物种的出生时间可以间接推断动物的死亡季节.但是由于分析得到的死亡年龄和动物的出生季节往往是分布区间,这种方法在判断动物死亡季节时精度较低[133]. ...
垩质年轮法在旧石器时代动物考古学中的应用
1
2014
... 动物考古学判断哺乳动物死亡年龄的方法,主要包括骨干骺端愈合情况、牙齿萌出次序和磨蚀程度、齿冠高度测量等[130].牙釉质坚硬易保存,因此经常通过牙齿萌出顺序和磨蚀程度来判断动物的死亡年龄,结合该物种的出生时间可以间接推断动物的死亡季节.但是由于分析得到的死亡年龄和动物的出生季节往往是分布区间,这种方法在判断动物死亡季节时精度较低[133]. ...
Histological observations of cementum growth in horse teeth and their application to archaeology
1
1995
... 牙骨质(cementum)是牙根周围的同心状增长层,为围绕牙周韧带的胶原纤维束矿化形成,厚约30~50 μm,含60%的骨胶原,40%的羟基磷灰石和少量非骨胶原蛋白质,一般分布于牙龈线以下的牙颈部及牙根外围.而奇蹄类动物如马的颊齿,其牙骨质不仅覆盖于牙根表面,还分布于除嚼面外的整颗牙齿外围.组织学研究结果表明,牙骨质细胞生长与个体食物摄取、所处的气候环境有关:当外界气候较为温暖、食物资源较为充足时,垩质细胞生长速度快且分布密集;反之气候干冷、营养匮乏时,垩质细胞生长速度缓慢同时分布稀疏[134].通过对山羊进行的现代控制实验发现,造成牙骨质条带生长呈现明暗变化的原因主要有两个,一是咀嚼不同食物的频率和强度改变了牙骨质中蛋白质束的方向;二是摄入营养的改变造成牙骨质条带矿化沉积相对速率的改变.两个原因都与动物的季节性食性变化相关,共同作用形成牙骨质条带生长呈现明暗变化[135].牙骨质从牙齿萌出开始终生生长,无重改造和重吸收作用,在偏振光下,哺乳动物牙骨质切片中的不透明条带表示生长速率缓慢的季节如冬天(也可以很宽),半透明条带表示持续快速的生长.反射光下切片的透明度与之相反,即透射光下的不透明条带为反射光的半透明条带,透射光下的透明条带为反射光的不透明条带[136](图4a).通过条带计数,加上该牙齿的萌出时间可以推断动物的死亡年龄;通过评估最外层的条带的类型,可以判断动物的死亡季节. ...
The biological basis for seasonal increments in dental cementum and their application to archaeological research
1
1994
... 牙骨质(cementum)是牙根周围的同心状增长层,为围绕牙周韧带的胶原纤维束矿化形成,厚约30~50 μm,含60%的骨胶原,40%的羟基磷灰石和少量非骨胶原蛋白质,一般分布于牙龈线以下的牙颈部及牙根外围.而奇蹄类动物如马的颊齿,其牙骨质不仅覆盖于牙根表面,还分布于除嚼面外的整颗牙齿外围.组织学研究结果表明,牙骨质细胞生长与个体食物摄取、所处的气候环境有关:当外界气候较为温暖、食物资源较为充足时,垩质细胞生长速度快且分布密集;反之气候干冷、营养匮乏时,垩质细胞生长速度缓慢同时分布稀疏[134].通过对山羊进行的现代控制实验发现,造成牙骨质条带生长呈现明暗变化的原因主要有两个,一是咀嚼不同食物的频率和强度改变了牙骨质中蛋白质束的方向;二是摄入营养的改变造成牙骨质条带矿化沉积相对速率的改变.两个原因都与动物的季节性食性变化相关,共同作用形成牙骨质条带生长呈现明暗变化[135].牙骨质从牙齿萌出开始终生生长,无重改造和重吸收作用,在偏振光下,哺乳动物牙骨质切片中的不透明条带表示生长速率缓慢的季节如冬天(也可以很宽),半透明条带表示持续快速的生长.反射光下切片的透明度与之相反,即透射光下的不透明条带为反射光的半透明条带,透射光下的透明条带为反射光的不透明条带[136](图4a).通过条带计数,加上该牙齿的萌出时间可以推断动物的死亡年龄;通过评估最外层的条带的类型,可以判断动物的死亡季节. ...
Seasonality of Gravettian sites in the Middle Danube Region and adjoining areas of Central Europe
2
2013
... 牙骨质(cementum)是牙根周围的同心状增长层,为围绕牙周韧带的胶原纤维束矿化形成,厚约30~50 μm,含60%的骨胶原,40%的羟基磷灰石和少量非骨胶原蛋白质,一般分布于牙龈线以下的牙颈部及牙根外围.而奇蹄类动物如马的颊齿,其牙骨质不仅覆盖于牙根表面,还分布于除嚼面外的整颗牙齿外围.组织学研究结果表明,牙骨质细胞生长与个体食物摄取、所处的气候环境有关:当外界气候较为温暖、食物资源较为充足时,垩质细胞生长速度快且分布密集;反之气候干冷、营养匮乏时,垩质细胞生长速度缓慢同时分布稀疏[134].通过对山羊进行的现代控制实验发现,造成牙骨质条带生长呈现明暗变化的原因主要有两个,一是咀嚼不同食物的频率和强度改变了牙骨质中蛋白质束的方向;二是摄入营养的改变造成牙骨质条带矿化沉积相对速率的改变.两个原因都与动物的季节性食性变化相关,共同作用形成牙骨质条带生长呈现明暗变化[135].牙骨质从牙齿萌出开始终生生长,无重改造和重吸收作用,在偏振光下,哺乳动物牙骨质切片中的不透明条带表示生长速率缓慢的季节如冬天(也可以很宽),半透明条带表示持续快速的生长.反射光下切片的透明度与之相反,即透射光下的不透明条带为反射光的半透明条带,透射光下的透明条带为反射光的不透明条带[136](图4a).通过条带计数,加上该牙齿的萌出时间可以推断动物的死亡年龄;通过评估最外层的条带的类型,可以判断动物的死亡季节. ...
... 注:a. 牙骨质位置和偏光显微镜下显微结构[136];b. 鱼脊椎[143];c. 鱼耳石[144-145];d. 鱼鳞;e. 鱼鳃盖骨[143,146];f. 鱼匙骨[147]. ...
A new method of age determination for mammals
1
1952
... 对牙骨质周期性沉积特征的研究始于20世纪50年代,代表性工作是Laws通过计算年轮数量来估算象海豹(Mirounga leonina)的死亡年龄的研究[137].20世纪60年代末,牙骨质年轮分析法被运用于考古遗址出土的动物化石研究中[138],通过比较现生绵羊与铁器时代绵羊的牙齿,推断铁器时代的绵羊死亡于夏季.而后,这种方法被推广到世界各地,涉及的动物主要包括鹿科、羊亚科和马科等,对讨论古代先民的捕食策略、遗址使用的季节和定居形式等问题提供了很大帮助[138⇓⇓-141].该方法在中国的考古遗址中的应用较少,目前仅见对许家窑遗址早、晚更新世地层出土的马牙进行过类似分析[142]. ...
Identification and interpretation of growth rings in the secondary dental cementum of Ovis aries L
2
1968
... 对牙骨质周期性沉积特征的研究始于20世纪50年代,代表性工作是Laws通过计算年轮数量来估算象海豹(Mirounga leonina)的死亡年龄的研究[137].20世纪60年代末,牙骨质年轮分析法被运用于考古遗址出土的动物化石研究中[138],通过比较现生绵羊与铁器时代绵羊的牙齿,推断铁器时代的绵羊死亡于夏季.而后,这种方法被推广到世界各地,涉及的动物主要包括鹿科、羊亚科和马科等,对讨论古代先民的捕食策略、遗址使用的季节和定居形式等问题提供了很大帮助[138⇓⇓-141].该方法在中国的考古遗址中的应用较少,目前仅见对许家窑遗址早、晚更新世地层出土的马牙进行过类似分析[142]. ...
... [138⇓⇓-141].该方法在中国的考古遗址中的应用较少,目前仅见对许家窑遗址早、晚更新世地层出土的马牙进行过类似分析[142]. ...
Using gazelle dental cementum studies to explore seasonality and mobility patterns of the Early-Middle Epipalaeolithic Azraq Basin, Jordan
1
2012
... 对牙骨质周期性沉积特征的研究始于20世纪50年代,代表性工作是Laws通过计算年轮数量来估算象海豹(Mirounga leonina)的死亡年龄的研究[137].20世纪60年代末,牙骨质年轮分析法被运用于考古遗址出土的动物化石研究中[138],通过比较现生绵羊与铁器时代绵羊的牙齿,推断铁器时代的绵羊死亡于夏季.而后,这种方法被推广到世界各地,涉及的动物主要包括鹿科、羊亚科和马科等,对讨论古代先民的捕食策略、遗址使用的季节和定居形式等问题提供了很大帮助[138⇓⇓-141].该方法在中国的考古遗址中的应用较少,目前仅见对许家窑遗址早、晚更新世地层出土的马牙进行过类似分析[142]. ...
Variability in seasonal mobility patterns in Bronze and Iron Age Kazakhstan through cementum analysis
1
2020
... 对牙骨质周期性沉积特征的研究始于20世纪50年代,代表性工作是Laws通过计算年轮数量来估算象海豹(Mirounga leonina)的死亡年龄的研究[137].20世纪60年代末,牙骨质年轮分析法被运用于考古遗址出土的动物化石研究中[138],通过比较现生绵羊与铁器时代绵羊的牙齿,推断铁器时代的绵羊死亡于夏季.而后,这种方法被推广到世界各地,涉及的动物主要包括鹿科、羊亚科和马科等,对讨论古代先民的捕食策略、遗址使用的季节和定居形式等问题提供了很大帮助[138⇓⇓-141].该方法在中国的考古遗址中的应用较少,目前仅见对许家窑遗址早、晚更新世地层出土的马牙进行过类似分析[142]. ...
Seasonal investigation of fallow deer cellular cementum from Mediterranean ecosystems and its implications for reliable cementochronology applications
1
2022
... 对牙骨质周期性沉积特征的研究始于20世纪50年代,代表性工作是Laws通过计算年轮数量来估算象海豹(Mirounga leonina)的死亡年龄的研究[137].20世纪60年代末,牙骨质年轮分析法被运用于考古遗址出土的动物化石研究中[138],通过比较现生绵羊与铁器时代绵羊的牙齿,推断铁器时代的绵羊死亡于夏季.而后,这种方法被推广到世界各地,涉及的动物主要包括鹿科、羊亚科和马科等,对讨论古代先民的捕食策略、遗址使用的季节和定居形式等问题提供了很大帮助[138⇓⇓-141].该方法在中国的考古遗址中的应用较少,目前仅见对许家窑遗址早、晚更新世地层出土的马牙进行过类似分析[142]. ...
A preliminary application of dental cementum incremental analysis to determine the season-of-death of equids from the Xujiayao site, China
1
2017
... 对牙骨质周期性沉积特征的研究始于20世纪50年代,代表性工作是Laws通过计算年轮数量来估算象海豹(Mirounga leonina)的死亡年龄的研究[137].20世纪60年代末,牙骨质年轮分析法被运用于考古遗址出土的动物化石研究中[138],通过比较现生绵羊与铁器时代绵羊的牙齿,推断铁器时代的绵羊死亡于夏季.而后,这种方法被推广到世界各地,涉及的动物主要包括鹿科、羊亚科和马科等,对讨论古代先民的捕食策略、遗址使用的季节和定居形式等问题提供了很大帮助[138⇓⇓-141].该方法在中国的考古遗址中的应用较少,目前仅见对许家窑遗址早、晚更新世地层出土的马牙进行过类似分析[142]. ...
Precision of age estimations from otolith, vertebra, and opercular bone of Gymnocypris firmispinatus (Actinopterygii: Cypriniformes: Cyprinidae) in the Anning River, China
2
2017
... 注:a. 牙骨质位置和偏光显微镜下显微结构[136];b. 鱼脊椎[143];c. 鱼耳石[144-145];d. 鱼鳞;e. 鱼鳃盖骨[143,146];f. 鱼匙骨[147]. ...
... [143,146];f. 鱼匙骨[147]. ...
Reconstructing age distribution, season of capture and growth rate of fish from archaeological sites based on otoliths and vertebrae
4
1999
... 注:a. 牙骨质位置和偏光显微镜下显微结构[136];b. 鱼脊椎[143];c. 鱼耳石[144-145];d. 鱼鳞;e. 鱼鳃盖骨[143,146];f. 鱼匙骨[147]. ...
... 鱼是变温动物,具有明显的周期性生长特性;鳞片、骨骼以及耳石等钙化结构终生生长,没有后期的改造和重吸收作用[150].通常认为,鱼类在冬季生长受限,形成狭窄的生长带,在透射光下呈半透明状,在反射光下相对较暗;相对较宽的区域(夏环)形成于快速生长时期,在透射光下呈不透明状,在反射光下呈白色;一个夏季环和一个冬季环即构成一个年轮,一般一年形成一个年轮,因此年轮的数量代表鱼的年龄[144,150].影响鱼类钙化结构年轮形成的机制尚未得到统一认识,通常认为与温度的季节性变化相关[151].但同时一些其他的因素如内部节律(繁殖、性别逆转、迁徙和性成熟)和外部条件(食物或水的丰匮、水质和盐度等)也与鱼类钙化结构年轮的形成具有一定的相关性[152].而对于热带鱼类,则可能更多地受到旱季和雨季交替的影响[153].在年轮形成规律的基础上,现代鱼类学的研究中常用边缘增长分析来判断鱼类年龄和研究鱼类生长,通常使用鱼鳞和耳石,原因在于鱼鳞在现代鱼类中最易获取,而耳石具有最高的精度和准确性,另外脊椎、匙骨、方骨、鳃盖骨和鳍棘等也比较常用(图4b~4f).边缘增长分析中,也可以通过测量边缘年轮与次外圈年轮的比值来反映鱼类的生长,即边缘增长率(Marginal Increment Ratio),可以通过按月测量提供一整年的边缘年轮增长曲线.但此方法难以准确测量微小增量,在生长缓慢和年龄较大的个体中存在较大误差[154]. ...
... 边缘增长分析也被广泛应用于动物考古学中,通过观察边缘年轮的生长情况来判断鱼类遗存的死亡季节[144,153,155 -156].这项研究的基础仍然是对目标物种的现生标本年轮形成季节的了解.由于年轮的形成受到鱼的种类、性别、采集地点和水体特征等多种因素的影响[152,155],因此需要对遗址所在区域的鱼类区系和资源状况,以及相关现生鱼类的个体生长情况,尤其是耳石和脊椎骨等的年轮生长情况进行详细的研究,才能为考古遗址出土鱼类的死亡年龄和季节提供确切的判断依据[144,153,156]. ...
... [144,153,156]. ...
Estimates on age, growth, and mortality of Gymnocypris firmispinatus (Cyprinidae: Schizothoracinae) in the Anning River, China
1
2019
... 注:a. 牙骨质位置和偏光显微镜下显微结构[136];b. 鱼脊椎[143];c. 鱼耳石[144-145];d. 鱼鳞;e. 鱼鳃盖骨[143,146];f. 鱼匙骨[147]. ...
Precision of age estimations from otolith, vertebra, and opercular bone of Gymnocypris firmispinatus (Actinopterygii: Cypriniformes: Cyprinidae) in the Anning River, China
1
2017
... 注:a. 牙骨质位置和偏光显微镜下显微结构[136];b. 鱼脊椎[143];c. 鱼耳石[144-145];d. 鱼鳞;e. 鱼鳃盖骨[143,146];f. 鱼匙骨[147]. ...
Precision and bias of cleithra and sagittal otoliths used to estimate ages of northern Pike
1
2013
... 注:a. 牙骨质位置和偏光显微镜下显微结构[136];b. 鱼脊椎[143];c. 鱼耳石[144-145];d. 鱼鳞;e. 鱼鳃盖骨[143,146];f. 鱼匙骨[147]. ...
Dental annuli age determination on white-tailed deer from archaeological sites
1
1974
... 限制牙骨质生长分析方法在考古学中广泛应用的一大重要因素是该方法的破坏性采样,因此,在制作牙骨质切片之前要首先进行动物考古学分析,并保存照片、甚至三维扫描模型等信息.此外,制作组织切片需要一整套较完备的硬组织切片设备,才能将误差控制到最小,并且其制作过程相对较复杂,最好有专业的人员进行操作才能保证结果的准确性.Kay借鉴了地质学家制作脆弱岩石切片的方法来制作动物化石切片,能更清楚地呈现垩质年轮;利用偏光显微镜观察,进一步分辨埋藏作用形成的假生长年轮,提高动物死亡季节判断的准确性[148-149]. ...
Polarizing microscopy identification of chemical diagenesis in archaeological cementum
1
2002
... 限制牙骨质生长分析方法在考古学中广泛应用的一大重要因素是该方法的破坏性采样,因此,在制作牙骨质切片之前要首先进行动物考古学分析,并保存照片、甚至三维扫描模型等信息.此外,制作组织切片需要一整套较完备的硬组织切片设备,才能将误差控制到最小,并且其制作过程相对较复杂,最好有专业的人员进行操作才能保证结果的准确性.Kay借鉴了地质学家制作脆弱岩石切片的方法来制作动物化石切片,能更清楚地呈现垩质年轮;利用偏光显微镜观察,进一步分辨埋藏作用形成的假生长年轮,提高动物死亡季节判断的准确性[148-149]. ...
The Ageing of Fish:The Proceedings of an International Symposium
2
1974
... 鱼是变温动物,具有明显的周期性生长特性;鳞片、骨骼以及耳石等钙化结构终生生长,没有后期的改造和重吸收作用[150].通常认为,鱼类在冬季生长受限,形成狭窄的生长带,在透射光下呈半透明状,在反射光下相对较暗;相对较宽的区域(夏环)形成于快速生长时期,在透射光下呈不透明状,在反射光下呈白色;一个夏季环和一个冬季环即构成一个年轮,一般一年形成一个年轮,因此年轮的数量代表鱼的年龄[144,150].影响鱼类钙化结构年轮形成的机制尚未得到统一认识,通常认为与温度的季节性变化相关[151].但同时一些其他的因素如内部节律(繁殖、性别逆转、迁徙和性成熟)和外部条件(食物或水的丰匮、水质和盐度等)也与鱼类钙化结构年轮的形成具有一定的相关性[152].而对于热带鱼类,则可能更多地受到旱季和雨季交替的影响[153].在年轮形成规律的基础上,现代鱼类学的研究中常用边缘增长分析来判断鱼类年龄和研究鱼类生长,通常使用鱼鳞和耳石,原因在于鱼鳞在现代鱼类中最易获取,而耳石具有最高的精度和准确性,另外脊椎、匙骨、方骨、鳃盖骨和鳍棘等也比较常用(图4b~4f).边缘增长分析中,也可以通过测量边缘年轮与次外圈年轮的比值来反映鱼类的生长,即边缘增长率(Marginal Increment Ratio),可以通过按月测量提供一整年的边缘年轮增长曲线.但此方法难以准确测量微小增量,在生长缓慢和年龄较大的个体中存在较大误差[154]. ...
... ,150].影响鱼类钙化结构年轮形成的机制尚未得到统一认识,通常认为与温度的季节性变化相关[151].但同时一些其他的因素如内部节律(繁殖、性别逆转、迁徙和性成熟)和外部条件(食物或水的丰匮、水质和盐度等)也与鱼类钙化结构年轮的形成具有一定的相关性[152].而对于热带鱼类,则可能更多地受到旱季和雨季交替的影响[153].在年轮形成规律的基础上,现代鱼类学的研究中常用边缘增长分析来判断鱼类年龄和研究鱼类生长,通常使用鱼鳞和耳石,原因在于鱼鳞在现代鱼类中最易获取,而耳石具有最高的精度和准确性,另外脊椎、匙骨、方骨、鳃盖骨和鳍棘等也比较常用(图4b~4f).边缘增长分析中,也可以通过测量边缘年轮与次外圈年轮的比值来反映鱼类的生长,即边缘增长率(Marginal Increment Ratio),可以通过按月测量提供一整年的边缘年轮增长曲线.但此方法难以准确测量微小增量,在生长缓慢和年龄较大的个体中存在较大误差[154]. ...
Shell midden sclerochronology
1
2011
... 鱼是变温动物,具有明显的周期性生长特性;鳞片、骨骼以及耳石等钙化结构终生生长,没有后期的改造和重吸收作用[150].通常认为,鱼类在冬季生长受限,形成狭窄的生长带,在透射光下呈半透明状,在反射光下相对较暗;相对较宽的区域(夏环)形成于快速生长时期,在透射光下呈不透明状,在反射光下呈白色;一个夏季环和一个冬季环即构成一个年轮,一般一年形成一个年轮,因此年轮的数量代表鱼的年龄[144,150].影响鱼类钙化结构年轮形成的机制尚未得到统一认识,通常认为与温度的季节性变化相关[151].但同时一些其他的因素如内部节律(繁殖、性别逆转、迁徙和性成熟)和外部条件(食物或水的丰匮、水质和盐度等)也与鱼类钙化结构年轮的形成具有一定的相关性[152].而对于热带鱼类,则可能更多地受到旱季和雨季交替的影响[153].在年轮形成规律的基础上,现代鱼类学的研究中常用边缘增长分析来判断鱼类年龄和研究鱼类生长,通常使用鱼鳞和耳石,原因在于鱼鳞在现代鱼类中最易获取,而耳石具有最高的精度和准确性,另外脊椎、匙骨、方骨、鳃盖骨和鳍棘等也比较常用(图4b~4f).边缘增长分析中,也可以通过测量边缘年轮与次外圈年轮的比值来反映鱼类的生长,即边缘增长率(Marginal Increment Ratio),可以通过按月测量提供一整年的边缘年轮增长曲线.但此方法难以准确测量微小增量,在生长缓慢和年龄较大的个体中存在较大误差[154]. ...
Handbook of fish age estimation protocols and validation methods
2
2019
... 鱼是变温动物,具有明显的周期性生长特性;鳞片、骨骼以及耳石等钙化结构终生生长,没有后期的改造和重吸收作用[150].通常认为,鱼类在冬季生长受限,形成狭窄的生长带,在透射光下呈半透明状,在反射光下相对较暗;相对较宽的区域(夏环)形成于快速生长时期,在透射光下呈不透明状,在反射光下呈白色;一个夏季环和一个冬季环即构成一个年轮,一般一年形成一个年轮,因此年轮的数量代表鱼的年龄[144,150].影响鱼类钙化结构年轮形成的机制尚未得到统一认识,通常认为与温度的季节性变化相关[151].但同时一些其他的因素如内部节律(繁殖、性别逆转、迁徙和性成熟)和外部条件(食物或水的丰匮、水质和盐度等)也与鱼类钙化结构年轮的形成具有一定的相关性[152].而对于热带鱼类,则可能更多地受到旱季和雨季交替的影响[153].在年轮形成规律的基础上,现代鱼类学的研究中常用边缘增长分析来判断鱼类年龄和研究鱼类生长,通常使用鱼鳞和耳石,原因在于鱼鳞在现代鱼类中最易获取,而耳石具有最高的精度和准确性,另外脊椎、匙骨、方骨、鳃盖骨和鳍棘等也比较常用(图4b~4f).边缘增长分析中,也可以通过测量边缘年轮与次外圈年轮的比值来反映鱼类的生长,即边缘增长率(Marginal Increment Ratio),可以通过按月测量提供一整年的边缘年轮增长曲线.但此方法难以准确测量微小增量,在生长缓慢和年龄较大的个体中存在较大误差[154]. ...
... 边缘增长分析也被广泛应用于动物考古学中,通过观察边缘年轮的生长情况来判断鱼类遗存的死亡季节[144,153,155 -156].这项研究的基础仍然是对目标物种的现生标本年轮形成季节的了解.由于年轮的形成受到鱼的种类、性别、采集地点和水体特征等多种因素的影响[152,155],因此需要对遗址所在区域的鱼类区系和资源状况,以及相关现生鱼类的个体生长情况,尤其是耳石和脊椎骨等的年轮生长情况进行详细的研究,才能为考古遗址出土鱼类的死亡年龄和季节提供确切的判断依据[144,153,156]. ...
Reconstructing freshwater fishing seasonality in a neotropical savanna: First application of swamp eel (Synbranchus marmoratus) sclerochronology to a pre-Columbian Amazonian site (Loma Salvatierra Bolivia)
3
2021
... 鱼是变温动物,具有明显的周期性生长特性;鳞片、骨骼以及耳石等钙化结构终生生长,没有后期的改造和重吸收作用[150].通常认为,鱼类在冬季生长受限,形成狭窄的生长带,在透射光下呈半透明状,在反射光下相对较暗;相对较宽的区域(夏环)形成于快速生长时期,在透射光下呈不透明状,在反射光下呈白色;一个夏季环和一个冬季环即构成一个年轮,一般一年形成一个年轮,因此年轮的数量代表鱼的年龄[144,150].影响鱼类钙化结构年轮形成的机制尚未得到统一认识,通常认为与温度的季节性变化相关[151].但同时一些其他的因素如内部节律(繁殖、性别逆转、迁徙和性成熟)和外部条件(食物或水的丰匮、水质和盐度等)也与鱼类钙化结构年轮的形成具有一定的相关性[152].而对于热带鱼类,则可能更多地受到旱季和雨季交替的影响[153].在年轮形成规律的基础上,现代鱼类学的研究中常用边缘增长分析来判断鱼类年龄和研究鱼类生长,通常使用鱼鳞和耳石,原因在于鱼鳞在现代鱼类中最易获取,而耳石具有最高的精度和准确性,另外脊椎、匙骨、方骨、鳃盖骨和鳍棘等也比较常用(图4b~4f).边缘增长分析中,也可以通过测量边缘年轮与次外圈年轮的比值来反映鱼类的生长,即边缘增长率(Marginal Increment Ratio),可以通过按月测量提供一整年的边缘年轮增长曲线.但此方法难以准确测量微小增量,在生长缓慢和年龄较大的个体中存在较大误差[154]. ...
... 边缘增长分析也被广泛应用于动物考古学中,通过观察边缘年轮的生长情况来判断鱼类遗存的死亡季节[144,153,155 -156].这项研究的基础仍然是对目标物种的现生标本年轮形成季节的了解.由于年轮的形成受到鱼的种类、性别、采集地点和水体特征等多种因素的影响[152,155],因此需要对遗址所在区域的鱼类区系和资源状况,以及相关现生鱼类的个体生长情况,尤其是耳石和脊椎骨等的年轮生长情况进行详细的研究,才能为考古遗址出土鱼类的死亡年龄和季节提供确切的判断依据[144,153,156]. ...
... ,153,156]. ...
Age validation of lemon sole (Microstomus kitt), using marginal increment analysis
1
2014
... 鱼是变温动物,具有明显的周期性生长特性;鳞片、骨骼以及耳石等钙化结构终生生长,没有后期的改造和重吸收作用[150].通常认为,鱼类在冬季生长受限,形成狭窄的生长带,在透射光下呈半透明状,在反射光下相对较暗;相对较宽的区域(夏环)形成于快速生长时期,在透射光下呈不透明状,在反射光下呈白色;一个夏季环和一个冬季环即构成一个年轮,一般一年形成一个年轮,因此年轮的数量代表鱼的年龄[144,150].影响鱼类钙化结构年轮形成的机制尚未得到统一认识,通常认为与温度的季节性变化相关[151].但同时一些其他的因素如内部节律(繁殖、性别逆转、迁徙和性成熟)和外部条件(食物或水的丰匮、水质和盐度等)也与鱼类钙化结构年轮的形成具有一定的相关性[152].而对于热带鱼类,则可能更多地受到旱季和雨季交替的影响[153].在年轮形成规律的基础上,现代鱼类学的研究中常用边缘增长分析来判断鱼类年龄和研究鱼类生长,通常使用鱼鳞和耳石,原因在于鱼鳞在现代鱼类中最易获取,而耳石具有最高的精度和准确性,另外脊椎、匙骨、方骨、鳃盖骨和鳍棘等也比较常用(图4b~4f).边缘增长分析中,也可以通过测量边缘年轮与次外圈年轮的比值来反映鱼类的生长,即边缘增长率(Marginal Increment Ratio),可以通过按月测量提供一整年的边缘年轮增长曲线.但此方法难以准确测量微小增量,在生长缓慢和年龄较大的个体中存在较大误差[154]. ...
Some archaeological uses of fish remains
2
1972
... 边缘增长分析也被广泛应用于动物考古学中,通过观察边缘年轮的生长情况来判断鱼类遗存的死亡季节[144,153,155 -156].这项研究的基础仍然是对目标物种的现生标本年轮形成季节的了解.由于年轮的形成受到鱼的种类、性别、采集地点和水体特征等多种因素的影响[152,155],因此需要对遗址所在区域的鱼类区系和资源状况,以及相关现生鱼类的个体生长情况,尤其是耳石和脊椎骨等的年轮生长情况进行详细的研究,才能为考古遗址出土鱼类的死亡年龄和季节提供确切的判断依据[144,153,156]. ...
... ,155],因此需要对遗址所在区域的鱼类区系和资源状况,以及相关现生鱼类的个体生长情况,尤其是耳石和脊椎骨等的年轮生长情况进行详细的研究,才能为考古遗址出土鱼类的死亡年龄和季节提供确切的判断依据[144,153,156]. ...
Seasonality of fishing in south-east Queensland
2
1992
... 边缘增长分析也被广泛应用于动物考古学中,通过观察边缘年轮的生长情况来判断鱼类遗存的死亡季节[144,153,155 -156].这项研究的基础仍然是对目标物种的现生标本年轮形成季节的了解.由于年轮的形成受到鱼的种类、性别、采集地点和水体特征等多种因素的影响[152,155],因此需要对遗址所在区域的鱼类区系和资源状况,以及相关现生鱼类的个体生长情况,尤其是耳石和脊椎骨等的年轮生长情况进行详细的研究,才能为考古遗址出土鱼类的死亡年龄和季节提供确切的判断依据[144,153,156]. ...
... ,156]. ...
羊卓雍错高原裸鲤(Gymnocypris waddellii Regan)繁殖生物学研究
3
2011
... 以雅鲁藏布江为例,分布于雅鲁藏布江流域中游干流和主要支流的现生鱼类共计25种,隶属于3目、5科、17属,其中高原鱼类的土著区系16种[157].主要经济鱼类有拉萨裸裂尻(Schizopygopsis younghusbandi)、异齿裂腹鱼(Schizothorax o'connori)、拉萨裂腹鱼(Racoma waltoni)、巨须裂腹鱼(Schizothorax macropogon)、双须叶须鱼(Ptychobarbus dipogon)、尖裸鲤(Oxygymnocypris stewartii)、黑斑原鮡(Glyptosternum maculatum)共7种,占渔获物重量的99%以上.其中拉萨裸裂尻、异齿裂腹鱼、巨须裂腹鱼3种鱼类占渔获物的82%[158-159].另外,高原裸鲤(Gymnocypris waddellii)大量分布于雅鲁藏布流域附近的羊卓雍错[157]和多庆错等地(表3). ...
... [157]和多庆错等地(表3). ...
... The formation time of winter-summer boundary of main fish in the middle reaches of the Yarlung Zangbo River
Tab. 3 | 物种 | 拉丁名 | 冬夏轮交界形成 | 判断
材料 | 参考文献 |
| 周期(a) | 时间(月份) |
| 双须叶须鱼 | Ptychobarbus dipogon | 1 | 3—5 | 耳石 | [166] |
| | 1 | 3—6 | 耳石 | [161] |
| 尖裸鲤 | Oxygymnocypris stewartii | 1 | 3—6
3—5 | 耳石
脊椎骨 | [160] |
| 拉萨裸裂尻 | Schizopygopsis younghusbandi | 1 | 3—5 | 耳石 | [167] |
| 黑斑原鮡 | Glyptosternum maculatum | 1 | 4—6 | 脊椎骨 | [162] |
| 异齿裂腹鱼 | Schizothorax o'connori | 1 | 3—5
3—6 | 耳石
脊椎骨 | [163] |
| 拉萨裂腹鱼 | Schizothorax waltoni | 1 | 3—5 | 耳石和
脊椎骨 | [164-165] |
| 巨须裂腹鱼 | Schizothorax macropogon | 1 | 3—6 | 耳石 | [168] |
| 高原裸鲤 | Gymnocypris waddellii | 1 | -- | 臀麟和
背鳍条 | [157] |
以上所提及雅鲁藏布江中游主要经济鱼类均有个体生物学研究发表,尤其对利用脊椎骨、耳石的年轮判断生长季节进行了较为详细的论述.对青藏高原本土鱼类的年龄与生长研究表明,利用耳石和脊椎骨判断动物年龄具有较高的精确性和可靠性,脊椎骨的准确性仅次于耳石[160-161].表3对雅鲁藏布江中游主要鱼类的耳石和脊椎骨冬轮与夏轮交界处形成周期和时间进行了总结.研究发现,尖裸鲤、黑斑原鮡、异齿裂腹鱼和拉萨裂腹鱼脊椎骨年轮的形成周期均为1年,且除黑斑原鮡的年轮形成于4—6月份外,其他3种鱼年轮的形成时间均为3—5月份[160,162⇓⇓ -165].双须叶须鱼、拉萨裸裂尻和巨须裂腹鱼耳石的年轮均形成于3—5月份或3—6月份[161,166⇓ -168],高原裸鲤暂无各骨骼年轮的形成时间数据.据此我们认为,可以通过对青藏高原上考古遗址出土鱼类脊椎骨年轮边缘类型的分析,确定史前鱼类的死亡季节,从而指示史前人类在该遗址的活动季节. ...
羊卓雍错高原裸鲤(Gymnocypris waddellii Regan)繁殖生物学研究
3
2011
... 以雅鲁藏布江为例,分布于雅鲁藏布江流域中游干流和主要支流的现生鱼类共计25种,隶属于3目、5科、17属,其中高原鱼类的土著区系16种[157].主要经济鱼类有拉萨裸裂尻(Schizopygopsis younghusbandi)、异齿裂腹鱼(Schizothorax o'connori)、拉萨裂腹鱼(Racoma waltoni)、巨须裂腹鱼(Schizothorax macropogon)、双须叶须鱼(Ptychobarbus dipogon)、尖裸鲤(Oxygymnocypris stewartii)、黑斑原鮡(Glyptosternum maculatum)共7种,占渔获物重量的99%以上.其中拉萨裸裂尻、异齿裂腹鱼、巨须裂腹鱼3种鱼类占渔获物的82%[158-159].另外,高原裸鲤(Gymnocypris waddellii)大量分布于雅鲁藏布流域附近的羊卓雍错[157]和多庆错等地(表3). ...
... [157]和多庆错等地(表3). ...
... The formation time of winter-summer boundary of main fish in the middle reaches of the Yarlung Zangbo River
Tab. 3 | 物种 | 拉丁名 | 冬夏轮交界形成 | 判断
材料 | 参考文献 |
| 周期(a) | 时间(月份) |
| 双须叶须鱼 | Ptychobarbus dipogon | 1 | 3—5 | 耳石 | [166] |
| | 1 | 3—6 | 耳石 | [161] |
| 尖裸鲤 | Oxygymnocypris stewartii | 1 | 3—6
3—5 | 耳石
脊椎骨 | [160] |
| 拉萨裸裂尻 | Schizopygopsis younghusbandi | 1 | 3—5 | 耳石 | [167] |
| 黑斑原鮡 | Glyptosternum maculatum | 1 | 4—6 | 脊椎骨 | [162] |
| 异齿裂腹鱼 | Schizothorax o'connori | 1 | 3—5
3—6 | 耳石
脊椎骨 | [163] |
| 拉萨裂腹鱼 | Schizothorax waltoni | 1 | 3—5 | 耳石和
脊椎骨 | [164-165] |
| 巨须裂腹鱼 | Schizothorax macropogon | 1 | 3—6 | 耳石 | [168] |
| 高原裸鲤 | Gymnocypris waddellii | 1 | -- | 臀麟和
背鳍条 | [157] |
以上所提及雅鲁藏布江中游主要经济鱼类均有个体生物学研究发表,尤其对利用脊椎骨、耳石的年轮判断生长季节进行了较为详细的论述.对青藏高原本土鱼类的年龄与生长研究表明,利用耳石和脊椎骨判断动物年龄具有较高的精确性和可靠性,脊椎骨的准确性仅次于耳石[160-161].表3对雅鲁藏布江中游主要鱼类的耳石和脊椎骨冬轮与夏轮交界处形成周期和时间进行了总结.研究发现,尖裸鲤、黑斑原鮡、异齿裂腹鱼和拉萨裂腹鱼脊椎骨年轮的形成周期均为1年,且除黑斑原鮡的年轮形成于4—6月份外,其他3种鱼年轮的形成时间均为3—5月份[160,162⇓⇓ -165].双须叶须鱼、拉萨裸裂尻和巨须裂腹鱼耳石的年轮均形成于3—5月份或3—6月份[161,166⇓ -168],高原裸鲤暂无各骨骼年轮的形成时间数据.据此我们认为,可以通过对青藏高原上考古遗址出土鱼类脊椎骨年轮边缘类型的分析,确定史前鱼类的死亡季节,从而指示史前人类在该遗址的活动季节. ...
雅鲁藏布江中游渔业资源现状研究
1
2010
... 以雅鲁藏布江为例,分布于雅鲁藏布江流域中游干流和主要支流的现生鱼类共计25种,隶属于3目、5科、17属,其中高原鱼类的土著区系16种[157].主要经济鱼类有拉萨裸裂尻(Schizopygopsis younghusbandi)、异齿裂腹鱼(Schizothorax o'connori)、拉萨裂腹鱼(Racoma waltoni)、巨须裂腹鱼(Schizothorax macropogon)、双须叶须鱼(Ptychobarbus dipogon)、尖裸鲤(Oxygymnocypris stewartii)、黑斑原鮡(Glyptosternum maculatum)共7种,占渔获物重量的99%以上.其中拉萨裸裂尻、异齿裂腹鱼、巨须裂腹鱼3种鱼类占渔获物的82%[158-159].另外,高原裸鲤(Gymnocypris waddellii)大量分布于雅鲁藏布流域附近的羊卓雍错[157]和多庆错等地(表3). ...
雅鲁藏布江中游渔业资源现状研究
1
2010
... 以雅鲁藏布江为例,分布于雅鲁藏布江流域中游干流和主要支流的现生鱼类共计25种,隶属于3目、5科、17属,其中高原鱼类的土著区系16种[157].主要经济鱼类有拉萨裸裂尻(Schizopygopsis younghusbandi)、异齿裂腹鱼(Schizothorax o'connori)、拉萨裂腹鱼(Racoma waltoni)、巨须裂腹鱼(Schizothorax macropogon)、双须叶须鱼(Ptychobarbus dipogon)、尖裸鲤(Oxygymnocypris stewartii)、黑斑原鮡(Glyptosternum maculatum)共7种,占渔获物重量的99%以上.其中拉萨裸裂尻、异齿裂腹鱼、巨须裂腹鱼3种鱼类占渔获物的82%[158-159].另外,高原裸鲤(Gymnocypris waddellii)大量分布于雅鲁藏布流域附近的羊卓雍错[157]和多庆错等地(表3). ...
雅鲁藏布江中上游鱼类区系和资源状况初步调查
1
2011
... 以雅鲁藏布江为例,分布于雅鲁藏布江流域中游干流和主要支流的现生鱼类共计25种,隶属于3目、5科、17属,其中高原鱼类的土著区系16种[157].主要经济鱼类有拉萨裸裂尻(Schizopygopsis younghusbandi)、异齿裂腹鱼(Schizothorax o'connori)、拉萨裂腹鱼(Racoma waltoni)、巨须裂腹鱼(Schizothorax macropogon)、双须叶须鱼(Ptychobarbus dipogon)、尖裸鲤(Oxygymnocypris stewartii)、黑斑原鮡(Glyptosternum maculatum)共7种,占渔获物重量的99%以上.其中拉萨裸裂尻、异齿裂腹鱼、巨须裂腹鱼3种鱼类占渔获物的82%[158-159].另外,高原裸鲤(Gymnocypris waddellii)大量分布于雅鲁藏布流域附近的羊卓雍错[157]和多庆错等地(表3). ...
雅鲁藏布江中上游鱼类区系和资源状况初步调查
1
2011
... 以雅鲁藏布江为例,分布于雅鲁藏布江流域中游干流和主要支流的现生鱼类共计25种,隶属于3目、5科、17属,其中高原鱼类的土著区系16种[157].主要经济鱼类有拉萨裸裂尻(Schizopygopsis younghusbandi)、异齿裂腹鱼(Schizothorax o'connori)、拉萨裂腹鱼(Racoma waltoni)、巨须裂腹鱼(Schizothorax macropogon)、双须叶须鱼(Ptychobarbus dipogon)、尖裸鲤(Oxygymnocypris stewartii)、黑斑原鮡(Glyptosternum maculatum)共7种,占渔获物重量的99%以上.其中拉萨裸裂尻、异齿裂腹鱼、巨须裂腹鱼3种鱼类占渔获物的82%[158-159].另外,高原裸鲤(Gymnocypris waddellii)大量分布于雅鲁藏布流域附近的羊卓雍错[157]和多庆错等地(表3). ...
3
2014
... The formation time of winter-summer boundary of main fish in the middle reaches of the Yarlung Zangbo River
Tab. 3 | 物种 | 拉丁名 | 冬夏轮交界形成 | 判断
材料 | 参考文献 |
| 周期(a) | 时间(月份) |
| 双须叶须鱼 | Ptychobarbus dipogon | 1 | 3—5 | 耳石 | [166] |
| | 1 | 3—6 | 耳石 | [161] |
| 尖裸鲤 | Oxygymnocypris stewartii | 1 | 3—6
3—5 | 耳石
脊椎骨 | [160] |
| 拉萨裸裂尻 | Schizopygopsis younghusbandi | 1 | 3—5 | 耳石 | [167] |
| 黑斑原鮡 | Glyptosternum maculatum | 1 | 4—6 | 脊椎骨 | [162] |
| 异齿裂腹鱼 | Schizothorax o'connori | 1 | 3—5
3—6 | 耳石
脊椎骨 | [163] |
| 拉萨裂腹鱼 | Schizothorax waltoni | 1 | 3—5 | 耳石和
脊椎骨 | [164-165] |
| 巨须裂腹鱼 | Schizothorax macropogon | 1 | 3—6 | 耳石 | [168] |
| 高原裸鲤 | Gymnocypris waddellii | 1 | -- | 臀麟和
背鳍条 | [157] |
以上所提及雅鲁藏布江中游主要经济鱼类均有个体生物学研究发表,尤其对利用脊椎骨、耳石的年轮判断生长季节进行了较为详细的论述.对青藏高原本土鱼类的年龄与生长研究表明,利用耳石和脊椎骨判断动物年龄具有较高的精确性和可靠性,脊椎骨的准确性仅次于耳石[160-161].表3对雅鲁藏布江中游主要鱼类的耳石和脊椎骨冬轮与夏轮交界处形成周期和时间进行了总结.研究发现,尖裸鲤、黑斑原鮡、异齿裂腹鱼和拉萨裂腹鱼脊椎骨年轮的形成周期均为1年,且除黑斑原鮡的年轮形成于4—6月份外,其他3种鱼年轮的形成时间均为3—5月份[160,162⇓⇓ -165].双须叶须鱼、拉萨裸裂尻和巨须裂腹鱼耳石的年轮均形成于3—5月份或3—6月份[161,166⇓ -168],高原裸鲤暂无各骨骼年轮的形成时间数据.据此我们认为,可以通过对青藏高原上考古遗址出土鱼类脊椎骨年轮边缘类型的分析,确定史前鱼类的死亡季节,从而指示史前人类在该遗址的活动季节. ...
... 以上所提及雅鲁藏布江中游主要经济鱼类均有个体生物学研究发表,尤其对利用脊椎骨、耳石的年轮判断生长季节进行了较为详细的论述.对青藏高原本土鱼类的年龄与生长研究表明,利用耳石和脊椎骨判断动物年龄具有较高的精确性和可靠性,脊椎骨的准确性仅次于耳石[160-161].表3对雅鲁藏布江中游主要鱼类的耳石和脊椎骨冬轮与夏轮交界处形成周期和时间进行了总结.研究发现,尖裸鲤、黑斑原鮡、异齿裂腹鱼和拉萨裂腹鱼脊椎骨年轮的形成周期均为1年,且除黑斑原鮡的年轮形成于4—6月份外,其他3种鱼年轮的形成时间均为3—5月份[160,162⇓⇓ -165].双须叶须鱼、拉萨裸裂尻和巨须裂腹鱼耳石的年轮均形成于3—5月份或3—6月份[161,166⇓ -168],高原裸鲤暂无各骨骼年轮的形成时间数据.据此我们认为,可以通过对青藏高原上考古遗址出土鱼类脊椎骨年轮边缘类型的分析,确定史前鱼类的死亡季节,从而指示史前人类在该遗址的活动季节. ...
... [160,162⇓⇓ -165].双须叶须鱼、拉萨裸裂尻和巨须裂腹鱼耳石的年轮均形成于3—5月份或3—6月份[161,166⇓ -168],高原裸鲤暂无各骨骼年轮的形成时间数据.据此我们认为,可以通过对青藏高原上考古遗址出土鱼类脊椎骨年轮边缘类型的分析,确定史前鱼类的死亡季节,从而指示史前人类在该遗址的活动季节. ...
3
2014
... The formation time of winter-summer boundary of main fish in the middle reaches of the Yarlung Zangbo River
Tab. 3 | 物种 | 拉丁名 | 冬夏轮交界形成 | 判断
材料 | 参考文献 |
| 周期(a) | 时间(月份) |
| 双须叶须鱼 | Ptychobarbus dipogon | 1 | 3—5 | 耳石 | [166] |
| | 1 | 3—6 | 耳石 | [161] |
| 尖裸鲤 | Oxygymnocypris stewartii | 1 | 3—6
3—5 | 耳石
脊椎骨 | [160] |
| 拉萨裸裂尻 | Schizopygopsis younghusbandi | 1 | 3—5 | 耳石 | [167] |
| 黑斑原鮡 | Glyptosternum maculatum | 1 | 4—6 | 脊椎骨 | [162] |
| 异齿裂腹鱼 | Schizothorax o'connori | 1 | 3—5
3—6 | 耳石
脊椎骨 | [163] |
| 拉萨裂腹鱼 | Schizothorax waltoni | 1 | 3—5 | 耳石和
脊椎骨 | [164-165] |
| 巨须裂腹鱼 | Schizothorax macropogon | 1 | 3—6 | 耳石 | [168] |
| 高原裸鲤 | Gymnocypris waddellii | 1 | -- | 臀麟和
背鳍条 | [157] |
以上所提及雅鲁藏布江中游主要经济鱼类均有个体生物学研究发表,尤其对利用脊椎骨、耳石的年轮判断生长季节进行了较为详细的论述.对青藏高原本土鱼类的年龄与生长研究表明,利用耳石和脊椎骨判断动物年龄具有较高的精确性和可靠性,脊椎骨的准确性仅次于耳石[160-161].表3对雅鲁藏布江中游主要鱼类的耳石和脊椎骨冬轮与夏轮交界处形成周期和时间进行了总结.研究发现,尖裸鲤、黑斑原鮡、异齿裂腹鱼和拉萨裂腹鱼脊椎骨年轮的形成周期均为1年,且除黑斑原鮡的年轮形成于4—6月份外,其他3种鱼年轮的形成时间均为3—5月份[160,162⇓⇓ -165].双须叶须鱼、拉萨裸裂尻和巨须裂腹鱼耳石的年轮均形成于3—5月份或3—6月份[161,166⇓ -168],高原裸鲤暂无各骨骼年轮的形成时间数据.据此我们认为,可以通过对青藏高原上考古遗址出土鱼类脊椎骨年轮边缘类型的分析,确定史前鱼类的死亡季节,从而指示史前人类在该遗址的活动季节. ...
... 以上所提及雅鲁藏布江中游主要经济鱼类均有个体生物学研究发表,尤其对利用脊椎骨、耳石的年轮判断生长季节进行了较为详细的论述.对青藏高原本土鱼类的年龄与生长研究表明,利用耳石和脊椎骨判断动物年龄具有较高的精确性和可靠性,脊椎骨的准确性仅次于耳石[160-161].表3对雅鲁藏布江中游主要鱼类的耳石和脊椎骨冬轮与夏轮交界处形成周期和时间进行了总结.研究发现,尖裸鲤、黑斑原鮡、异齿裂腹鱼和拉萨裂腹鱼脊椎骨年轮的形成周期均为1年,且除黑斑原鮡的年轮形成于4—6月份外,其他3种鱼年轮的形成时间均为3—5月份[160,162⇓⇓ -165].双须叶须鱼、拉萨裸裂尻和巨须裂腹鱼耳石的年轮均形成于3—5月份或3—6月份[161,166⇓ -168],高原裸鲤暂无各骨骼年轮的形成时间数据.据此我们认为,可以通过对青藏高原上考古遗址出土鱼类脊椎骨年轮边缘类型的分析,确定史前鱼类的死亡季节,从而指示史前人类在该遗址的活动季节. ...
... [160,162⇓⇓ -165].双须叶须鱼、拉萨裸裂尻和巨须裂腹鱼耳石的年轮均形成于3—5月份或3—6月份[161,166⇓ -168],高原裸鲤暂无各骨骼年轮的形成时间数据.据此我们认为,可以通过对青藏高原上考古遗址出土鱼类脊椎骨年轮边缘类型的分析,确定史前鱼类的死亡季节,从而指示史前人类在该遗址的活动季节. ...
3
2015
... The formation time of winter-summer boundary of main fish in the middle reaches of the Yarlung Zangbo River
Tab. 3 | 物种 | 拉丁名 | 冬夏轮交界形成 | 判断
材料 | 参考文献 |
| 周期(a) | 时间(月份) |
| 双须叶须鱼 | Ptychobarbus dipogon | 1 | 3—5 | 耳石 | [166] |
| | 1 | 3—6 | 耳石 | [161] |
| 尖裸鲤 | Oxygymnocypris stewartii | 1 | 3—6
3—5 | 耳石
脊椎骨 | [160] |
| 拉萨裸裂尻 | Schizopygopsis younghusbandi | 1 | 3—5 | 耳石 | [167] |
| 黑斑原鮡 | Glyptosternum maculatum | 1 | 4—6 | 脊椎骨 | [162] |
| 异齿裂腹鱼 | Schizothorax o'connori | 1 | 3—5
3—6 | 耳石
脊椎骨 | [163] |
| 拉萨裂腹鱼 | Schizothorax waltoni | 1 | 3—5 | 耳石和
脊椎骨 | [164-165] |
| 巨须裂腹鱼 | Schizothorax macropogon | 1 | 3—6 | 耳石 | [168] |
| 高原裸鲤 | Gymnocypris waddellii | 1 | -- | 臀麟和
背鳍条 | [157] |
以上所提及雅鲁藏布江中游主要经济鱼类均有个体生物学研究发表,尤其对利用脊椎骨、耳石的年轮判断生长季节进行了较为详细的论述.对青藏高原本土鱼类的年龄与生长研究表明,利用耳石和脊椎骨判断动物年龄具有较高的精确性和可靠性,脊椎骨的准确性仅次于耳石[160-161].表3对雅鲁藏布江中游主要鱼类的耳石和脊椎骨冬轮与夏轮交界处形成周期和时间进行了总结.研究发现,尖裸鲤、黑斑原鮡、异齿裂腹鱼和拉萨裂腹鱼脊椎骨年轮的形成周期均为1年,且除黑斑原鮡的年轮形成于4—6月份外,其他3种鱼年轮的形成时间均为3—5月份[160,162⇓⇓ -165].双须叶须鱼、拉萨裸裂尻和巨须裂腹鱼耳石的年轮均形成于3—5月份或3—6月份[161,166⇓ -168],高原裸鲤暂无各骨骼年轮的形成时间数据.据此我们认为,可以通过对青藏高原上考古遗址出土鱼类脊椎骨年轮边缘类型的分析,确定史前鱼类的死亡季节,从而指示史前人类在该遗址的活动季节. ...
... 以上所提及雅鲁藏布江中游主要经济鱼类均有个体生物学研究发表,尤其对利用脊椎骨、耳石的年轮判断生长季节进行了较为详细的论述.对青藏高原本土鱼类的年龄与生长研究表明,利用耳石和脊椎骨判断动物年龄具有较高的精确性和可靠性,脊椎骨的准确性仅次于耳石[160-161].表3对雅鲁藏布江中游主要鱼类的耳石和脊椎骨冬轮与夏轮交界处形成周期和时间进行了总结.研究发现,尖裸鲤、黑斑原鮡、异齿裂腹鱼和拉萨裂腹鱼脊椎骨年轮的形成周期均为1年,且除黑斑原鮡的年轮形成于4—6月份外,其他3种鱼年轮的形成时间均为3—5月份[160,162⇓⇓ -165].双须叶须鱼、拉萨裸裂尻和巨须裂腹鱼耳石的年轮均形成于3—5月份或3—6月份[161,166⇓ -168],高原裸鲤暂无各骨骼年轮的形成时间数据.据此我们认为,可以通过对青藏高原上考古遗址出土鱼类脊椎骨年轮边缘类型的分析,确定史前鱼类的死亡季节,从而指示史前人类在该遗址的活动季节. ...
... [161,166⇓ -168],高原裸鲤暂无各骨骼年轮的形成时间数据.据此我们认为,可以通过对青藏高原上考古遗址出土鱼类脊椎骨年轮边缘类型的分析,确定史前鱼类的死亡季节,从而指示史前人类在该遗址的活动季节. ...
3
2015
... The formation time of winter-summer boundary of main fish in the middle reaches of the Yarlung Zangbo River
Tab. 3 | 物种 | 拉丁名 | 冬夏轮交界形成 | 判断
材料 | 参考文献 |
| 周期(a) | 时间(月份) |
| 双须叶须鱼 | Ptychobarbus dipogon | 1 | 3—5 | 耳石 | [166] |
| | 1 | 3—6 | 耳石 | [161] |
| 尖裸鲤 | Oxygymnocypris stewartii | 1 | 3—6
3—5 | 耳石
脊椎骨 | [160] |
| 拉萨裸裂尻 | Schizopygopsis younghusbandi | 1 | 3—5 | 耳石 | [167] |
| 黑斑原鮡 | Glyptosternum maculatum | 1 | 4—6 | 脊椎骨 | [162] |
| 异齿裂腹鱼 | Schizothorax o'connori | 1 | 3—5
3—6 | 耳石
脊椎骨 | [163] |
| 拉萨裂腹鱼 | Schizothorax waltoni | 1 | 3—5 | 耳石和
脊椎骨 | [164-165] |
| 巨须裂腹鱼 | Schizothorax macropogon | 1 | 3—6 | 耳石 | [168] |
| 高原裸鲤 | Gymnocypris waddellii | 1 | -- | 臀麟和
背鳍条 | [157] |
以上所提及雅鲁藏布江中游主要经济鱼类均有个体生物学研究发表,尤其对利用脊椎骨、耳石的年轮判断生长季节进行了较为详细的论述.对青藏高原本土鱼类的年龄与生长研究表明,利用耳石和脊椎骨判断动物年龄具有较高的精确性和可靠性,脊椎骨的准确性仅次于耳石[160-161].表3对雅鲁藏布江中游主要鱼类的耳石和脊椎骨冬轮与夏轮交界处形成周期和时间进行了总结.研究发现,尖裸鲤、黑斑原鮡、异齿裂腹鱼和拉萨裂腹鱼脊椎骨年轮的形成周期均为1年,且除黑斑原鮡的年轮形成于4—6月份外,其他3种鱼年轮的形成时间均为3—5月份[160,162⇓⇓ -165].双须叶须鱼、拉萨裸裂尻和巨须裂腹鱼耳石的年轮均形成于3—5月份或3—6月份[161,166⇓ -168],高原裸鲤暂无各骨骼年轮的形成时间数据.据此我们认为,可以通过对青藏高原上考古遗址出土鱼类脊椎骨年轮边缘类型的分析,确定史前鱼类的死亡季节,从而指示史前人类在该遗址的活动季节. ...
... 以上所提及雅鲁藏布江中游主要经济鱼类均有个体生物学研究发表,尤其对利用脊椎骨、耳石的年轮判断生长季节进行了较为详细的论述.对青藏高原本土鱼类的年龄与生长研究表明,利用耳石和脊椎骨判断动物年龄具有较高的精确性和可靠性,脊椎骨的准确性仅次于耳石[160-161].表3对雅鲁藏布江中游主要鱼类的耳石和脊椎骨冬轮与夏轮交界处形成周期和时间进行了总结.研究发现,尖裸鲤、黑斑原鮡、异齿裂腹鱼和拉萨裂腹鱼脊椎骨年轮的形成周期均为1年,且除黑斑原鮡的年轮形成于4—6月份外,其他3种鱼年轮的形成时间均为3—5月份[160,162⇓⇓ -165].双须叶须鱼、拉萨裸裂尻和巨须裂腹鱼耳石的年轮均形成于3—5月份或3—6月份[161,166⇓ -168],高原裸鲤暂无各骨骼年轮的形成时间数据.据此我们认为,可以通过对青藏高原上考古遗址出土鱼类脊椎骨年轮边缘类型的分析,确定史前鱼类的死亡季节,从而指示史前人类在该遗址的活动季节. ...
... [161,166⇓ -168],高原裸鲤暂无各骨骼年轮的形成时间数据.据此我们认为,可以通过对青藏高原上考古遗址出土鱼类脊椎骨年轮边缘类型的分析,确定史前鱼类的死亡季节,从而指示史前人类在该遗址的活动季节. ...
雅鲁藏布江黑斑原鮡的年龄与生长
2
2008
... The formation time of winter-summer boundary of main fish in the middle reaches of the Yarlung Zangbo River
Tab. 3 | 物种 | 拉丁名 | 冬夏轮交界形成 | 判断
材料 | 参考文献 |
| 周期(a) | 时间(月份) |
| 双须叶须鱼 | Ptychobarbus dipogon | 1 | 3—5 | 耳石 | [166] |
| | 1 | 3—6 | 耳石 | [161] |
| 尖裸鲤 | Oxygymnocypris stewartii | 1 | 3—6
3—5 | 耳石
脊椎骨 | [160] |
| 拉萨裸裂尻 | Schizopygopsis younghusbandi | 1 | 3—5 | 耳石 | [167] |
| 黑斑原鮡 | Glyptosternum maculatum | 1 | 4—6 | 脊椎骨 | [162] |
| 异齿裂腹鱼 | Schizothorax o'connori | 1 | 3—5
3—6 | 耳石
脊椎骨 | [163] |
| 拉萨裂腹鱼 | Schizothorax waltoni | 1 | 3—5 | 耳石和
脊椎骨 | [164-165] |
| 巨须裂腹鱼 | Schizothorax macropogon | 1 | 3—6 | 耳石 | [168] |
| 高原裸鲤 | Gymnocypris waddellii | 1 | -- | 臀麟和
背鳍条 | [157] |
以上所提及雅鲁藏布江中游主要经济鱼类均有个体生物学研究发表,尤其对利用脊椎骨、耳石的年轮判断生长季节进行了较为详细的论述.对青藏高原本土鱼类的年龄与生长研究表明,利用耳石和脊椎骨判断动物年龄具有较高的精确性和可靠性,脊椎骨的准确性仅次于耳石[160-161].表3对雅鲁藏布江中游主要鱼类的耳石和脊椎骨冬轮与夏轮交界处形成周期和时间进行了总结.研究发现,尖裸鲤、黑斑原鮡、异齿裂腹鱼和拉萨裂腹鱼脊椎骨年轮的形成周期均为1年,且除黑斑原鮡的年轮形成于4—6月份外,其他3种鱼年轮的形成时间均为3—5月份[160,162⇓⇓ -165].双须叶须鱼、拉萨裸裂尻和巨须裂腹鱼耳石的年轮均形成于3—5月份或3—6月份[161,166⇓ -168],高原裸鲤暂无各骨骼年轮的形成时间数据.据此我们认为,可以通过对青藏高原上考古遗址出土鱼类脊椎骨年轮边缘类型的分析,确定史前鱼类的死亡季节,从而指示史前人类在该遗址的活动季节. ...
... 以上所提及雅鲁藏布江中游主要经济鱼类均有个体生物学研究发表,尤其对利用脊椎骨、耳石的年轮判断生长季节进行了较为详细的论述.对青藏高原本土鱼类的年龄与生长研究表明,利用耳石和脊椎骨判断动物年龄具有较高的精确性和可靠性,脊椎骨的准确性仅次于耳石[160-161].表3对雅鲁藏布江中游主要鱼类的耳石和脊椎骨冬轮与夏轮交界处形成周期和时间进行了总结.研究发现,尖裸鲤、黑斑原鮡、异齿裂腹鱼和拉萨裂腹鱼脊椎骨年轮的形成周期均为1年,且除黑斑原鮡的年轮形成于4—6月份外,其他3种鱼年轮的形成时间均为3—5月份[160,162⇓⇓ -165].双须叶须鱼、拉萨裸裂尻和巨须裂腹鱼耳石的年轮均形成于3—5月份或3—6月份[161,166⇓ -168],高原裸鲤暂无各骨骼年轮的形成时间数据.据此我们认为,可以通过对青藏高原上考古遗址出土鱼类脊椎骨年轮边缘类型的分析,确定史前鱼类的死亡季节,从而指示史前人类在该遗址的活动季节. ...
雅鲁藏布江黑斑原鮡的年龄与生长
2
2008
... The formation time of winter-summer boundary of main fish in the middle reaches of the Yarlung Zangbo River
Tab. 3 | 物种 | 拉丁名 | 冬夏轮交界形成 | 判断
材料 | 参考文献 |
| 周期(a) | 时间(月份) |
| 双须叶须鱼 | Ptychobarbus dipogon | 1 | 3—5 | 耳石 | [166] |
| | 1 | 3—6 | 耳石 | [161] |
| 尖裸鲤 | Oxygymnocypris stewartii | 1 | 3—6
3—5 | 耳石
脊椎骨 | [160] |
| 拉萨裸裂尻 | Schizopygopsis younghusbandi | 1 | 3—5 | 耳石 | [167] |
| 黑斑原鮡 | Glyptosternum maculatum | 1 | 4—6 | 脊椎骨 | [162] |
| 异齿裂腹鱼 | Schizothorax o'connori | 1 | 3—5
3—6 | 耳石
脊椎骨 | [163] |
| 拉萨裂腹鱼 | Schizothorax waltoni | 1 | 3—5 | 耳石和
脊椎骨 | [164-165] |
| 巨须裂腹鱼 | Schizothorax macropogon | 1 | 3—6 | 耳石 | [168] |
| 高原裸鲤 | Gymnocypris waddellii | 1 | -- | 臀麟和
背鳍条 | [157] |
以上所提及雅鲁藏布江中游主要经济鱼类均有个体生物学研究发表,尤其对利用脊椎骨、耳石的年轮判断生长季节进行了较为详细的论述.对青藏高原本土鱼类的年龄与生长研究表明,利用耳石和脊椎骨判断动物年龄具有较高的精确性和可靠性,脊椎骨的准确性仅次于耳石[160-161].表3对雅鲁藏布江中游主要鱼类的耳石和脊椎骨冬轮与夏轮交界处形成周期和时间进行了总结.研究发现,尖裸鲤、黑斑原鮡、异齿裂腹鱼和拉萨裂腹鱼脊椎骨年轮的形成周期均为1年,且除黑斑原鮡的年轮形成于4—6月份外,其他3种鱼年轮的形成时间均为3—5月份[160,162⇓⇓ -165].双须叶须鱼、拉萨裸裂尻和巨须裂腹鱼耳石的年轮均形成于3—5月份或3—6月份[161,166⇓ -168],高原裸鲤暂无各骨骼年轮的形成时间数据.据此我们认为,可以通过对青藏高原上考古遗址出土鱼类脊椎骨年轮边缘类型的分析,确定史前鱼类的死亡季节,从而指示史前人类在该遗址的活动季节. ...
... 以上所提及雅鲁藏布江中游主要经济鱼类均有个体生物学研究发表,尤其对利用脊椎骨、耳石的年轮判断生长季节进行了较为详细的论述.对青藏高原本土鱼类的年龄与生长研究表明,利用耳石和脊椎骨判断动物年龄具有较高的精确性和可靠性,脊椎骨的准确性仅次于耳石[160-161].表3对雅鲁藏布江中游主要鱼类的耳石和脊椎骨冬轮与夏轮交界处形成周期和时间进行了总结.研究发现,尖裸鲤、黑斑原鮡、异齿裂腹鱼和拉萨裂腹鱼脊椎骨年轮的形成周期均为1年,且除黑斑原鮡的年轮形成于4—6月份外,其他3种鱼年轮的形成时间均为3—5月份[160,162⇓⇓ -165].双须叶须鱼、拉萨裸裂尻和巨须裂腹鱼耳石的年轮均形成于3—5月份或3—6月份[161,166⇓ -168],高原裸鲤暂无各骨骼年轮的形成时间数据.据此我们认为,可以通过对青藏高原上考古遗址出土鱼类脊椎骨年轮边缘类型的分析,确定史前鱼类的死亡季节,从而指示史前人类在该遗址的活动季节. ...
Age validation, and comparison of otolith, vertebra and opercular bone for estimating age of Schizothorax o'connori in the Yarlung Tsangpo River, Tibet
2
2011
... The formation time of winter-summer boundary of main fish in the middle reaches of the Yarlung Zangbo River
Tab. 3 | 物种 | 拉丁名 | 冬夏轮交界形成 | 判断
材料 | 参考文献 |
| 周期(a) | 时间(月份) |
| 双须叶须鱼 | Ptychobarbus dipogon | 1 | 3—5 | 耳石 | [166] |
| | 1 | 3—6 | 耳石 | [161] |
| 尖裸鲤 | Oxygymnocypris stewartii | 1 | 3—6
3—5 | 耳石
脊椎骨 | [160] |
| 拉萨裸裂尻 | Schizopygopsis younghusbandi | 1 | 3—5 | 耳石 | [167] |
| 黑斑原鮡 | Glyptosternum maculatum | 1 | 4—6 | 脊椎骨 | [162] |
| 异齿裂腹鱼 | Schizothorax o'connori | 1 | 3—5
3—6 | 耳石
脊椎骨 | [163] |
| 拉萨裂腹鱼 | Schizothorax waltoni | 1 | 3—5 | 耳石和
脊椎骨 | [164-165] |
| 巨须裂腹鱼 | Schizothorax macropogon | 1 | 3—6 | 耳石 | [168] |
| 高原裸鲤 | Gymnocypris waddellii | 1 | -- | 臀麟和
背鳍条 | [157] |
以上所提及雅鲁藏布江中游主要经济鱼类均有个体生物学研究发表,尤其对利用脊椎骨、耳石的年轮判断生长季节进行了较为详细的论述.对青藏高原本土鱼类的年龄与生长研究表明,利用耳石和脊椎骨判断动物年龄具有较高的精确性和可靠性,脊椎骨的准确性仅次于耳石[160-161].表3对雅鲁藏布江中游主要鱼类的耳石和脊椎骨冬轮与夏轮交界处形成周期和时间进行了总结.研究发现,尖裸鲤、黑斑原鮡、异齿裂腹鱼和拉萨裂腹鱼脊椎骨年轮的形成周期均为1年,且除黑斑原鮡的年轮形成于4—6月份外,其他3种鱼年轮的形成时间均为3—5月份[160,162⇓⇓ -165].双须叶须鱼、拉萨裸裂尻和巨须裂腹鱼耳石的年轮均形成于3—5月份或3—6月份[161,166⇓ -168],高原裸鲤暂无各骨骼年轮的形成时间数据.据此我们认为,可以通过对青藏高原上考古遗址出土鱼类脊椎骨年轮边缘类型的分析,确定史前鱼类的死亡季节,从而指示史前人类在该遗址的活动季节. ...
... 以上所提及雅鲁藏布江中游主要经济鱼类均有个体生物学研究发表,尤其对利用脊椎骨、耳石的年轮判断生长季节进行了较为详细的论述.对青藏高原本土鱼类的年龄与生长研究表明,利用耳石和脊椎骨判断动物年龄具有较高的精确性和可靠性,脊椎骨的准确性仅次于耳石[160-161].表3对雅鲁藏布江中游主要鱼类的耳石和脊椎骨冬轮与夏轮交界处形成周期和时间进行了总结.研究发现,尖裸鲤、黑斑原鮡、异齿裂腹鱼和拉萨裂腹鱼脊椎骨年轮的形成周期均为1年,且除黑斑原鮡的年轮形成于4—6月份外,其他3种鱼年轮的形成时间均为3—5月份[160,162⇓⇓ -165].双须叶须鱼、拉萨裸裂尻和巨须裂腹鱼耳石的年轮均形成于3—5月份或3—6月份[161,166⇓ -168],高原裸鲤暂无各骨骼年轮的形成时间数据.据此我们认为,可以通过对青藏高原上考古遗址出土鱼类脊椎骨年轮边缘类型的分析,确定史前鱼类的死亡季节,从而指示史前人类在该遗址的活动季节. ...
2
2005
... The formation time of winter-summer boundary of main fish in the middle reaches of the Yarlung Zangbo River
Tab. 3 | 物种 | 拉丁名 | 冬夏轮交界形成 | 判断
材料 | 参考文献 |
| 周期(a) | 时间(月份) |
| 双须叶须鱼 | Ptychobarbus dipogon | 1 | 3—5 | 耳石 | [166] |
| | 1 | 3—6 | 耳石 | [161] |
| 尖裸鲤 | Oxygymnocypris stewartii | 1 | 3—6
3—5 | 耳石
脊椎骨 | [160] |
| 拉萨裸裂尻 | Schizopygopsis younghusbandi | 1 | 3—5 | 耳石 | [167] |
| 黑斑原鮡 | Glyptosternum maculatum | 1 | 4—6 | 脊椎骨 | [162] |
| 异齿裂腹鱼 | Schizothorax o'connori | 1 | 3—5
3—6 | 耳石
脊椎骨 | [163] |
| 拉萨裂腹鱼 | Schizothorax waltoni | 1 | 3—5 | 耳石和
脊椎骨 | [164-165] |
| 巨须裂腹鱼 | Schizothorax macropogon | 1 | 3—6 | 耳石 | [168] |
| 高原裸鲤 | Gymnocypris waddellii | 1 | -- | 臀麟和
背鳍条 | [157] |
以上所提及雅鲁藏布江中游主要经济鱼类均有个体生物学研究发表,尤其对利用脊椎骨、耳石的年轮判断生长季节进行了较为详细的论述.对青藏高原本土鱼类的年龄与生长研究表明,利用耳石和脊椎骨判断动物年龄具有较高的精确性和可靠性,脊椎骨的准确性仅次于耳石[160-161].表3对雅鲁藏布江中游主要鱼类的耳石和脊椎骨冬轮与夏轮交界处形成周期和时间进行了总结.研究发现,尖裸鲤、黑斑原鮡、异齿裂腹鱼和拉萨裂腹鱼脊椎骨年轮的形成周期均为1年,且除黑斑原鮡的年轮形成于4—6月份外,其他3种鱼年轮的形成时间均为3—5月份[160,162⇓⇓ -165].双须叶须鱼、拉萨裸裂尻和巨须裂腹鱼耳石的年轮均形成于3—5月份或3—6月份[161,166⇓ -168],高原裸鲤暂无各骨骼年轮的形成时间数据.据此我们认为,可以通过对青藏高原上考古遗址出土鱼类脊椎骨年轮边缘类型的分析,确定史前鱼类的死亡季节,从而指示史前人类在该遗址的活动季节. ...
... 以上所提及雅鲁藏布江中游主要经济鱼类均有个体生物学研究发表,尤其对利用脊椎骨、耳石的年轮判断生长季节进行了较为详细的论述.对青藏高原本土鱼类的年龄与生长研究表明,利用耳石和脊椎骨判断动物年龄具有较高的精确性和可靠性,脊椎骨的准确性仅次于耳石[160-161].表3对雅鲁藏布江中游主要鱼类的耳石和脊椎骨冬轮与夏轮交界处形成周期和时间进行了总结.研究发现,尖裸鲤、黑斑原鮡、异齿裂腹鱼和拉萨裂腹鱼脊椎骨年轮的形成周期均为1年,且除黑斑原鮡的年轮形成于4—6月份外,其他3种鱼年轮的形成时间均为3—5月份[160,162⇓⇓ -165].双须叶须鱼、拉萨裸裂尻和巨须裂腹鱼耳石的年轮均形成于3—5月份或3—6月份[161,166⇓ -168],高原裸鲤暂无各骨骼年轮的形成时间数据.据此我们认为,可以通过对青藏高原上考古遗址出土鱼类脊椎骨年轮边缘类型的分析,确定史前鱼类的死亡季节,从而指示史前人类在该遗址的活动季节. ...
2
2005
... The formation time of winter-summer boundary of main fish in the middle reaches of the Yarlung Zangbo River
Tab. 3 | 物种 | 拉丁名 | 冬夏轮交界形成 | 判断
材料 | 参考文献 |
| 周期(a) | 时间(月份) |
| 双须叶须鱼 | Ptychobarbus dipogon | 1 | 3—5 | 耳石 | [166] |
| | 1 | 3—6 | 耳石 | [161] |
| 尖裸鲤 | Oxygymnocypris stewartii | 1 | 3—6
3—5 | 耳石
脊椎骨 | [160] |
| 拉萨裸裂尻 | Schizopygopsis younghusbandi | 1 | 3—5 | 耳石 | [167] |
| 黑斑原鮡 | Glyptosternum maculatum | 1 | 4—6 | 脊椎骨 | [162] |
| 异齿裂腹鱼 | Schizothorax o'connori | 1 | 3—5
3—6 | 耳石
脊椎骨 | [163] |
| 拉萨裂腹鱼 | Schizothorax waltoni | 1 | 3—5 | 耳石和
脊椎骨 | [164-165] |
| 巨须裂腹鱼 | Schizothorax macropogon | 1 | 3—6 | 耳石 | [168] |
| 高原裸鲤 | Gymnocypris waddellii | 1 | -- | 臀麟和
背鳍条 | [157] |
以上所提及雅鲁藏布江中游主要经济鱼类均有个体生物学研究发表,尤其对利用脊椎骨、耳石的年轮判断生长季节进行了较为详细的论述.对青藏高原本土鱼类的年龄与生长研究表明,利用耳石和脊椎骨判断动物年龄具有较高的精确性和可靠性,脊椎骨的准确性仅次于耳石[160-161].表3对雅鲁藏布江中游主要鱼类的耳石和脊椎骨冬轮与夏轮交界处形成周期和时间进行了总结.研究发现,尖裸鲤、黑斑原鮡、异齿裂腹鱼和拉萨裂腹鱼脊椎骨年轮的形成周期均为1年,且除黑斑原鮡的年轮形成于4—6月份外,其他3种鱼年轮的形成时间均为3—5月份[160,162⇓⇓ -165].双须叶须鱼、拉萨裸裂尻和巨须裂腹鱼耳石的年轮均形成于3—5月份或3—6月份[161,166⇓ -168],高原裸鲤暂无各骨骼年轮的形成时间数据.据此我们认为,可以通过对青藏高原上考古遗址出土鱼类脊椎骨年轮边缘类型的分析,确定史前鱼类的死亡季节,从而指示史前人类在该遗址的活动季节. ...
... 以上所提及雅鲁藏布江中游主要经济鱼类均有个体生物学研究发表,尤其对利用脊椎骨、耳石的年轮判断生长季节进行了较为详细的论述.对青藏高原本土鱼类的年龄与生长研究表明,利用耳石和脊椎骨判断动物年龄具有较高的精确性和可靠性,脊椎骨的准确性仅次于耳石[160-161].表3对雅鲁藏布江中游主要鱼类的耳石和脊椎骨冬轮与夏轮交界处形成周期和时间进行了总结.研究发现,尖裸鲤、黑斑原鮡、异齿裂腹鱼和拉萨裂腹鱼脊椎骨年轮的形成周期均为1年,且除黑斑原鮡的年轮形成于4—6月份外,其他3种鱼年轮的形成时间均为3—5月份[160,162⇓⇓ -165].双须叶须鱼、拉萨裸裂尻和巨须裂腹鱼耳石的年轮均形成于3—5月份或3—6月份[161,166⇓ -168],高原裸鲤暂无各骨骼年轮的形成时间数据.据此我们认为,可以通过对青藏高原上考古遗址出土鱼类脊椎骨年轮边缘类型的分析,确定史前鱼类的死亡季节,从而指示史前人类在该遗址的活动季节. ...
2
2014
... The formation time of winter-summer boundary of main fish in the middle reaches of the Yarlung Zangbo River
Tab. 3 | 物种 | 拉丁名 | 冬夏轮交界形成 | 判断
材料 | 参考文献 |
| 周期(a) | 时间(月份) |
| 双须叶须鱼 | Ptychobarbus dipogon | 1 | 3—5 | 耳石 | [166] |
| | 1 | 3—6 | 耳石 | [161] |
| 尖裸鲤 | Oxygymnocypris stewartii | 1 | 3—6
3—5 | 耳石
脊椎骨 | [160] |
| 拉萨裸裂尻 | Schizopygopsis younghusbandi | 1 | 3—5 | 耳石 | [167] |
| 黑斑原鮡 | Glyptosternum maculatum | 1 | 4—6 | 脊椎骨 | [162] |
| 异齿裂腹鱼 | Schizothorax o'connori | 1 | 3—5
3—6 | 耳石
脊椎骨 | [163] |
| 拉萨裂腹鱼 | Schizothorax waltoni | 1 | 3—5 | 耳石和
脊椎骨 | [164-165] |
| 巨须裂腹鱼 | Schizothorax macropogon | 1 | 3—6 | 耳石 | [168] |
| 高原裸鲤 | Gymnocypris waddellii | 1 | -- | 臀麟和
背鳍条 | [157] |
以上所提及雅鲁藏布江中游主要经济鱼类均有个体生物学研究发表,尤其对利用脊椎骨、耳石的年轮判断生长季节进行了较为详细的论述.对青藏高原本土鱼类的年龄与生长研究表明,利用耳石和脊椎骨判断动物年龄具有较高的精确性和可靠性,脊椎骨的准确性仅次于耳石[160-161].表3对雅鲁藏布江中游主要鱼类的耳石和脊椎骨冬轮与夏轮交界处形成周期和时间进行了总结.研究发现,尖裸鲤、黑斑原鮡、异齿裂腹鱼和拉萨裂腹鱼脊椎骨年轮的形成周期均为1年,且除黑斑原鮡的年轮形成于4—6月份外,其他3种鱼年轮的形成时间均为3—5月份[160,162⇓⇓ -165].双须叶须鱼、拉萨裸裂尻和巨须裂腹鱼耳石的年轮均形成于3—5月份或3—6月份[161,166⇓ -168],高原裸鲤暂无各骨骼年轮的形成时间数据.据此我们认为,可以通过对青藏高原上考古遗址出土鱼类脊椎骨年轮边缘类型的分析,确定史前鱼类的死亡季节,从而指示史前人类在该遗址的活动季节. ...
... 以上所提及雅鲁藏布江中游主要经济鱼类均有个体生物学研究发表,尤其对利用脊椎骨、耳石的年轮判断生长季节进行了较为详细的论述.对青藏高原本土鱼类的年龄与生长研究表明,利用耳石和脊椎骨判断动物年龄具有较高的精确性和可靠性,脊椎骨的准确性仅次于耳石[160-161].表3对雅鲁藏布江中游主要鱼类的耳石和脊椎骨冬轮与夏轮交界处形成周期和时间进行了总结.研究发现,尖裸鲤、黑斑原鮡、异齿裂腹鱼和拉萨裂腹鱼脊椎骨年轮的形成周期均为1年,且除黑斑原鮡的年轮形成于4—6月份外,其他3种鱼年轮的形成时间均为3—5月份[160,162⇓⇓ -165].双须叶须鱼、拉萨裸裂尻和巨须裂腹鱼耳石的年轮均形成于3—5月份或3—6月份[161,166⇓ -168],高原裸鲤暂无各骨骼年轮的形成时间数据.据此我们认为,可以通过对青藏高原上考古遗址出土鱼类脊椎骨年轮边缘类型的分析,确定史前鱼类的死亡季节,从而指示史前人类在该遗址的活动季节. ...
2
2014
... The formation time of winter-summer boundary of main fish in the middle reaches of the Yarlung Zangbo River
Tab. 3 | 物种 | 拉丁名 | 冬夏轮交界形成 | 判断
材料 | 参考文献 |
| 周期(a) | 时间(月份) |
| 双须叶须鱼 | Ptychobarbus dipogon | 1 | 3—5 | 耳石 | [166] |
| | 1 | 3—6 | 耳石 | [161] |
| 尖裸鲤 | Oxygymnocypris stewartii | 1 | 3—6
3—5 | 耳石
脊椎骨 | [160] |
| 拉萨裸裂尻 | Schizopygopsis younghusbandi | 1 | 3—5 | 耳石 | [167] |
| 黑斑原鮡 | Glyptosternum maculatum | 1 | 4—6 | 脊椎骨 | [162] |
| 异齿裂腹鱼 | Schizothorax o'connori | 1 | 3—5
3—6 | 耳石
脊椎骨 | [163] |
| 拉萨裂腹鱼 | Schizothorax waltoni | 1 | 3—5 | 耳石和
脊椎骨 | [164-165] |
| 巨须裂腹鱼 | Schizothorax macropogon | 1 | 3—6 | 耳石 | [168] |
| 高原裸鲤 | Gymnocypris waddellii | 1 | -- | 臀麟和
背鳍条 | [157] |
以上所提及雅鲁藏布江中游主要经济鱼类均有个体生物学研究发表,尤其对利用脊椎骨、耳石的年轮判断生长季节进行了较为详细的论述.对青藏高原本土鱼类的年龄与生长研究表明,利用耳石和脊椎骨判断动物年龄具有较高的精确性和可靠性,脊椎骨的准确性仅次于耳石[160-161].表3对雅鲁藏布江中游主要鱼类的耳石和脊椎骨冬轮与夏轮交界处形成周期和时间进行了总结.研究发现,尖裸鲤、黑斑原鮡、异齿裂腹鱼和拉萨裂腹鱼脊椎骨年轮的形成周期均为1年,且除黑斑原鮡的年轮形成于4—6月份外,其他3种鱼年轮的形成时间均为3—5月份[160,162⇓⇓ -165].双须叶须鱼、拉萨裸裂尻和巨须裂腹鱼耳石的年轮均形成于3—5月份或3—6月份[161,166⇓ -168],高原裸鲤暂无各骨骼年轮的形成时间数据.据此我们认为,可以通过对青藏高原上考古遗址出土鱼类脊椎骨年轮边缘类型的分析,确定史前鱼类的死亡季节,从而指示史前人类在该遗址的活动季节. ...
... 以上所提及雅鲁藏布江中游主要经济鱼类均有个体生物学研究发表,尤其对利用脊椎骨、耳石的年轮判断生长季节进行了较为详细的论述.对青藏高原本土鱼类的年龄与生长研究表明,利用耳石和脊椎骨判断动物年龄具有较高的精确性和可靠性,脊椎骨的准确性仅次于耳石[160-161].表3对雅鲁藏布江中游主要鱼类的耳石和脊椎骨冬轮与夏轮交界处形成周期和时间进行了总结.研究发现,尖裸鲤、黑斑原鮡、异齿裂腹鱼和拉萨裂腹鱼脊椎骨年轮的形成周期均为1年,且除黑斑原鮡的年轮形成于4—6月份外,其他3种鱼年轮的形成时间均为3—5月份[160,162⇓⇓ -165].双须叶须鱼、拉萨裸裂尻和巨须裂腹鱼耳石的年轮均形成于3—5月份或3—6月份[161,166⇓ -168],高原裸鲤暂无各骨骼年轮的形成时间数据.据此我们认为,可以通过对青藏高原上考古遗址出土鱼类脊椎骨年轮边缘类型的分析,确定史前鱼类的死亡季节,从而指示史前人类在该遗址的活动季节. ...
Otolith characteristics and age determination of an endemic Ptychobarbus dipogon (Regan, 1905) (Cyprinidae: Schizothoracinae) in the Yarlung Tsangpo River, Tibet
2
2009
... The formation time of winter-summer boundary of main fish in the middle reaches of the Yarlung Zangbo River
Tab. 3 | 物种 | 拉丁名 | 冬夏轮交界形成 | 判断
材料 | 参考文献 |
| 周期(a) | 时间(月份) |
| 双须叶须鱼 | Ptychobarbus dipogon | 1 | 3—5 | 耳石 | [166] |
| | 1 | 3—6 | 耳石 | [161] |
| 尖裸鲤 | Oxygymnocypris stewartii | 1 | 3—6
3—5 | 耳石
脊椎骨 | [160] |
| 拉萨裸裂尻 | Schizopygopsis younghusbandi | 1 | 3—5 | 耳石 | [167] |
| 黑斑原鮡 | Glyptosternum maculatum | 1 | 4—6 | 脊椎骨 | [162] |
| 异齿裂腹鱼 | Schizothorax o'connori | 1 | 3—5
3—6 | 耳石
脊椎骨 | [163] |
| 拉萨裂腹鱼 | Schizothorax waltoni | 1 | 3—5 | 耳石和
脊椎骨 | [164-165] |
| 巨须裂腹鱼 | Schizothorax macropogon | 1 | 3—6 | 耳石 | [168] |
| 高原裸鲤 | Gymnocypris waddellii | 1 | -- | 臀麟和
背鳍条 | [157] |
以上所提及雅鲁藏布江中游主要经济鱼类均有个体生物学研究发表,尤其对利用脊椎骨、耳石的年轮判断生长季节进行了较为详细的论述.对青藏高原本土鱼类的年龄与生长研究表明,利用耳石和脊椎骨判断动物年龄具有较高的精确性和可靠性,脊椎骨的准确性仅次于耳石[160-161].表3对雅鲁藏布江中游主要鱼类的耳石和脊椎骨冬轮与夏轮交界处形成周期和时间进行了总结.研究发现,尖裸鲤、黑斑原鮡、异齿裂腹鱼和拉萨裂腹鱼脊椎骨年轮的形成周期均为1年,且除黑斑原鮡的年轮形成于4—6月份外,其他3种鱼年轮的形成时间均为3—5月份[160,162⇓⇓ -165].双须叶须鱼、拉萨裸裂尻和巨须裂腹鱼耳石的年轮均形成于3—5月份或3—6月份[161,166⇓ -168],高原裸鲤暂无各骨骼年轮的形成时间数据.据此我们认为,可以通过对青藏高原上考古遗址出土鱼类脊椎骨年轮边缘类型的分析,确定史前鱼类的死亡季节,从而指示史前人类在该遗址的活动季节. ...
... 以上所提及雅鲁藏布江中游主要经济鱼类均有个体生物学研究发表,尤其对利用脊椎骨、耳石的年轮判断生长季节进行了较为详细的论述.对青藏高原本土鱼类的年龄与生长研究表明,利用耳石和脊椎骨判断动物年龄具有较高的精确性和可靠性,脊椎骨的准确性仅次于耳石[160-161].表3对雅鲁藏布江中游主要鱼类的耳石和脊椎骨冬轮与夏轮交界处形成周期和时间进行了总结.研究发现,尖裸鲤、黑斑原鮡、异齿裂腹鱼和拉萨裂腹鱼脊椎骨年轮的形成周期均为1年,且除黑斑原鮡的年轮形成于4—6月份外,其他3种鱼年轮的形成时间均为3—5月份[160,162⇓⇓ -165].双须叶须鱼、拉萨裸裂尻和巨须裂腹鱼耳石的年轮均形成于3—5月份或3—6月份[161,166⇓ -168],高原裸鲤暂无各骨骼年轮的形成时间数据.据此我们认为,可以通过对青藏高原上考古遗址出土鱼类脊椎骨年轮边缘类型的分析,确定史前鱼类的死亡季节,从而指示史前人类在该遗址的活动季节. ...
2
2015
... The formation time of winter-summer boundary of main fish in the middle reaches of the Yarlung Zangbo River
Tab. 3 | 物种 | 拉丁名 | 冬夏轮交界形成 | 判断
材料 | 参考文献 |
| 周期(a) | 时间(月份) |
| 双须叶须鱼 | Ptychobarbus dipogon | 1 | 3—5 | 耳石 | [166] |
| | 1 | 3—6 | 耳石 | [161] |
| 尖裸鲤 | Oxygymnocypris stewartii | 1 | 3—6
3—5 | 耳石
脊椎骨 | [160] |
| 拉萨裸裂尻 | Schizopygopsis younghusbandi | 1 | 3—5 | 耳石 | [167] |
| 黑斑原鮡 | Glyptosternum maculatum | 1 | 4—6 | 脊椎骨 | [162] |
| 异齿裂腹鱼 | Schizothorax o'connori | 1 | 3—5
3—6 | 耳石
脊椎骨 | [163] |
| 拉萨裂腹鱼 | Schizothorax waltoni | 1 | 3—5 | 耳石和
脊椎骨 | [164-165] |
| 巨须裂腹鱼 | Schizothorax macropogon | 1 | 3—6 | 耳石 | [168] |
| 高原裸鲤 | Gymnocypris waddellii | 1 | -- | 臀麟和
背鳍条 | [157] |
以上所提及雅鲁藏布江中游主要经济鱼类均有个体生物学研究发表,尤其对利用脊椎骨、耳石的年轮判断生长季节进行了较为详细的论述.对青藏高原本土鱼类的年龄与生长研究表明,利用耳石和脊椎骨判断动物年龄具有较高的精确性和可靠性,脊椎骨的准确性仅次于耳石[160-161].表3对雅鲁藏布江中游主要鱼类的耳石和脊椎骨冬轮与夏轮交界处形成周期和时间进行了总结.研究发现,尖裸鲤、黑斑原鮡、异齿裂腹鱼和拉萨裂腹鱼脊椎骨年轮的形成周期均为1年,且除黑斑原鮡的年轮形成于4—6月份外,其他3种鱼年轮的形成时间均为3—5月份[160,162⇓⇓ -165].双须叶须鱼、拉萨裸裂尻和巨须裂腹鱼耳石的年轮均形成于3—5月份或3—6月份[161,166⇓ -168],高原裸鲤暂无各骨骼年轮的形成时间数据.据此我们认为,可以通过对青藏高原上考古遗址出土鱼类脊椎骨年轮边缘类型的分析,确定史前鱼类的死亡季节,从而指示史前人类在该遗址的活动季节. ...
... 以上所提及雅鲁藏布江中游主要经济鱼类均有个体生物学研究发表,尤其对利用脊椎骨、耳石的年轮判断生长季节进行了较为详细的论述.对青藏高原本土鱼类的年龄与生长研究表明,利用耳石和脊椎骨判断动物年龄具有较高的精确性和可靠性,脊椎骨的准确性仅次于耳石[160-161].表3对雅鲁藏布江中游主要鱼类的耳石和脊椎骨冬轮与夏轮交界处形成周期和时间进行了总结.研究发现,尖裸鲤、黑斑原鮡、异齿裂腹鱼和拉萨裂腹鱼脊椎骨年轮的形成周期均为1年,且除黑斑原鮡的年轮形成于4—6月份外,其他3种鱼年轮的形成时间均为3—5月份[160,162⇓⇓ -165].双须叶须鱼、拉萨裸裂尻和巨须裂腹鱼耳石的年轮均形成于3—5月份或3—6月份[161,166⇓ -168],高原裸鲤暂无各骨骼年轮的形成时间数据.据此我们认为,可以通过对青藏高原上考古遗址出土鱼类脊椎骨年轮边缘类型的分析,确定史前鱼类的死亡季节,从而指示史前人类在该遗址的活动季节. ...
2
2015
... The formation time of winter-summer boundary of main fish in the middle reaches of the Yarlung Zangbo River
Tab. 3 | 物种 | 拉丁名 | 冬夏轮交界形成 | 判断
材料 | 参考文献 |
| 周期(a) | 时间(月份) |
| 双须叶须鱼 | Ptychobarbus dipogon | 1 | 3—5 | 耳石 | [166] |
| | 1 | 3—6 | 耳石 | [161] |
| 尖裸鲤 | Oxygymnocypris stewartii | 1 | 3—6
3—5 | 耳石
脊椎骨 | [160] |
| 拉萨裸裂尻 | Schizopygopsis younghusbandi | 1 | 3—5 | 耳石 | [167] |
| 黑斑原鮡 | Glyptosternum maculatum | 1 | 4—6 | 脊椎骨 | [162] |
| 异齿裂腹鱼 | Schizothorax o'connori | 1 | 3—5
3—6 | 耳石
脊椎骨 | [163] |
| 拉萨裂腹鱼 | Schizothorax waltoni | 1 | 3—5 | 耳石和
脊椎骨 | [164-165] |
| 巨须裂腹鱼 | Schizothorax macropogon | 1 | 3—6 | 耳石 | [168] |
| 高原裸鲤 | Gymnocypris waddellii | 1 | -- | 臀麟和
背鳍条 | [157] |
以上所提及雅鲁藏布江中游主要经济鱼类均有个体生物学研究发表,尤其对利用脊椎骨、耳石的年轮判断生长季节进行了较为详细的论述.对青藏高原本土鱼类的年龄与生长研究表明,利用耳石和脊椎骨判断动物年龄具有较高的精确性和可靠性,脊椎骨的准确性仅次于耳石[160-161].表3对雅鲁藏布江中游主要鱼类的耳石和脊椎骨冬轮与夏轮交界处形成周期和时间进行了总结.研究发现,尖裸鲤、黑斑原鮡、异齿裂腹鱼和拉萨裂腹鱼脊椎骨年轮的形成周期均为1年,且除黑斑原鮡的年轮形成于4—6月份外,其他3种鱼年轮的形成时间均为3—5月份[160,162⇓⇓ -165].双须叶须鱼、拉萨裸裂尻和巨须裂腹鱼耳石的年轮均形成于3—5月份或3—6月份[161,166⇓ -168],高原裸鲤暂无各骨骼年轮的形成时间数据.据此我们认为,可以通过对青藏高原上考古遗址出土鱼类脊椎骨年轮边缘类型的分析,确定史前鱼类的死亡季节,从而指示史前人类在该遗址的活动季节. ...
... 以上所提及雅鲁藏布江中游主要经济鱼类均有个体生物学研究发表,尤其对利用脊椎骨、耳石的年轮判断生长季节进行了较为详细的论述.对青藏高原本土鱼类的年龄与生长研究表明,利用耳石和脊椎骨判断动物年龄具有较高的精确性和可靠性,脊椎骨的准确性仅次于耳石[160-161].表3对雅鲁藏布江中游主要鱼类的耳石和脊椎骨冬轮与夏轮交界处形成周期和时间进行了总结.研究发现,尖裸鲤、黑斑原鮡、异齿裂腹鱼和拉萨裂腹鱼脊椎骨年轮的形成周期均为1年,且除黑斑原鮡的年轮形成于4—6月份外,其他3种鱼年轮的形成时间均为3—5月份[160,162⇓⇓ -165].双须叶须鱼、拉萨裸裂尻和巨须裂腹鱼耳石的年轮均形成于3—5月份或3—6月份[161,166⇓ -168],高原裸鲤暂无各骨骼年轮的形成时间数据.据此我们认为,可以通过对青藏高原上考古遗址出土鱼类脊椎骨年轮边缘类型的分析,确定史前鱼类的死亡季节,从而指示史前人类在该遗址的活动季节. ...
2
2016
... The formation time of winter-summer boundary of main fish in the middle reaches of the Yarlung Zangbo River
Tab. 3 | 物种 | 拉丁名 | 冬夏轮交界形成 | 判断
材料 | 参考文献 |
| 周期(a) | 时间(月份) |
| 双须叶须鱼 | Ptychobarbus dipogon | 1 | 3—5 | 耳石 | [166] |
| | 1 | 3—6 | 耳石 | [161] |
| 尖裸鲤 | Oxygymnocypris stewartii | 1 | 3—6
3—5 | 耳石
脊椎骨 | [160] |
| 拉萨裸裂尻 | Schizopygopsis younghusbandi | 1 | 3—5 | 耳石 | [167] |
| 黑斑原鮡 | Glyptosternum maculatum | 1 | 4—6 | 脊椎骨 | [162] |
| 异齿裂腹鱼 | Schizothorax o'connori | 1 | 3—5
3—6 | 耳石
脊椎骨 | [163] |
| 拉萨裂腹鱼 | Schizothorax waltoni | 1 | 3—5 | 耳石和
脊椎骨 | [164-165] |
| 巨须裂腹鱼 | Schizothorax macropogon | 1 | 3—6 | 耳石 | [168] |
| 高原裸鲤 | Gymnocypris waddellii | 1 | -- | 臀麟和
背鳍条 | [157] |
以上所提及雅鲁藏布江中游主要经济鱼类均有个体生物学研究发表,尤其对利用脊椎骨、耳石的年轮判断生长季节进行了较为详细的论述.对青藏高原本土鱼类的年龄与生长研究表明,利用耳石和脊椎骨判断动物年龄具有较高的精确性和可靠性,脊椎骨的准确性仅次于耳石[160-161].表3对雅鲁藏布江中游主要鱼类的耳石和脊椎骨冬轮与夏轮交界处形成周期和时间进行了总结.研究发现,尖裸鲤、黑斑原鮡、异齿裂腹鱼和拉萨裂腹鱼脊椎骨年轮的形成周期均为1年,且除黑斑原鮡的年轮形成于4—6月份外,其他3种鱼年轮的形成时间均为3—5月份[160,162⇓⇓ -165].双须叶须鱼、拉萨裸裂尻和巨须裂腹鱼耳石的年轮均形成于3—5月份或3—6月份[161,166⇓ -168],高原裸鲤暂无各骨骼年轮的形成时间数据.据此我们认为,可以通过对青藏高原上考古遗址出土鱼类脊椎骨年轮边缘类型的分析,确定史前鱼类的死亡季节,从而指示史前人类在该遗址的活动季节. ...
... 以上所提及雅鲁藏布江中游主要经济鱼类均有个体生物学研究发表,尤其对利用脊椎骨、耳石的年轮判断生长季节进行了较为详细的论述.对青藏高原本土鱼类的年龄与生长研究表明,利用耳石和脊椎骨判断动物年龄具有较高的精确性和可靠性,脊椎骨的准确性仅次于耳石[160-161].表3对雅鲁藏布江中游主要鱼类的耳石和脊椎骨冬轮与夏轮交界处形成周期和时间进行了总结.研究发现,尖裸鲤、黑斑原鮡、异齿裂腹鱼和拉萨裂腹鱼脊椎骨年轮的形成周期均为1年,且除黑斑原鮡的年轮形成于4—6月份外,其他3种鱼年轮的形成时间均为3—5月份[160,162⇓⇓ -165].双须叶须鱼、拉萨裸裂尻和巨须裂腹鱼耳石的年轮均形成于3—5月份或3—6月份[161,166⇓ -168],高原裸鲤暂无各骨骼年轮的形成时间数据.据此我们认为,可以通过对青藏高原上考古遗址出土鱼类脊椎骨年轮边缘类型的分析,确定史前鱼类的死亡季节,从而指示史前人类在该遗址的活动季节. ...
2
2016
... The formation time of winter-summer boundary of main fish in the middle reaches of the Yarlung Zangbo River
Tab. 3 | 物种 | 拉丁名 | 冬夏轮交界形成 | 判断
材料 | 参考文献 |
| 周期(a) | 时间(月份) |
| 双须叶须鱼 | Ptychobarbus dipogon | 1 | 3—5 | 耳石 | [166] |
| | 1 | 3—6 | 耳石 | [161] |
| 尖裸鲤 | Oxygymnocypris stewartii | 1 | 3—6
3—5 | 耳石
脊椎骨 | [160] |
| 拉萨裸裂尻 | Schizopygopsis younghusbandi | 1 | 3—5 | 耳石 | [167] |
| 黑斑原鮡 | Glyptosternum maculatum | 1 | 4—6 | 脊椎骨 | [162] |
| 异齿裂腹鱼 | Schizothorax o'connori | 1 | 3—5
3—6 | 耳石
脊椎骨 | [163] |
| 拉萨裂腹鱼 | Schizothorax waltoni | 1 | 3—5 | 耳石和
脊椎骨 | [164-165] |
| 巨须裂腹鱼 | Schizothorax macropogon | 1 | 3—6 | 耳石 | [168] |
| 高原裸鲤 | Gymnocypris waddellii | 1 | -- | 臀麟和
背鳍条 | [157] |
以上所提及雅鲁藏布江中游主要经济鱼类均有个体生物学研究发表,尤其对利用脊椎骨、耳石的年轮判断生长季节进行了较为详细的论述.对青藏高原本土鱼类的年龄与生长研究表明,利用耳石和脊椎骨判断动物年龄具有较高的精确性和可靠性,脊椎骨的准确性仅次于耳石[160-161].表3对雅鲁藏布江中游主要鱼类的耳石和脊椎骨冬轮与夏轮交界处形成周期和时间进行了总结.研究发现,尖裸鲤、黑斑原鮡、异齿裂腹鱼和拉萨裂腹鱼脊椎骨年轮的形成周期均为1年,且除黑斑原鮡的年轮形成于4—6月份外,其他3种鱼年轮的形成时间均为3—5月份[160,162⇓⇓ -165].双须叶须鱼、拉萨裸裂尻和巨须裂腹鱼耳石的年轮均形成于3—5月份或3—6月份[161,166⇓ -168],高原裸鲤暂无各骨骼年轮的形成时间数据.据此我们认为,可以通过对青藏高原上考古遗址出土鱼类脊椎骨年轮边缘类型的分析,确定史前鱼类的死亡季节,从而指示史前人类在该遗址的活动季节. ...
... 以上所提及雅鲁藏布江中游主要经济鱼类均有个体生物学研究发表,尤其对利用脊椎骨、耳石的年轮判断生长季节进行了较为详细的论述.对青藏高原本土鱼类的年龄与生长研究表明,利用耳石和脊椎骨判断动物年龄具有较高的精确性和可靠性,脊椎骨的准确性仅次于耳石[160-161].表3对雅鲁藏布江中游主要鱼类的耳石和脊椎骨冬轮与夏轮交界处形成周期和时间进行了总结.研究发现,尖裸鲤、黑斑原鮡、异齿裂腹鱼和拉萨裂腹鱼脊椎骨年轮的形成周期均为1年,且除黑斑原鮡的年轮形成于4—6月份外,其他3种鱼年轮的形成时间均为3—5月份[160,162⇓⇓ -165].双须叶须鱼、拉萨裸裂尻和巨须裂腹鱼耳石的年轮均形成于3—5月份或3—6月份[161,166⇓ -168],高原裸鲤暂无各骨骼年轮的形成时间数据.据此我们认为,可以通过对青藏高原上考古遗址出土鱼类脊椎骨年轮边缘类型的分析,确定史前鱼类的死亡季节,从而指示史前人类在该遗址的活动季节. ...


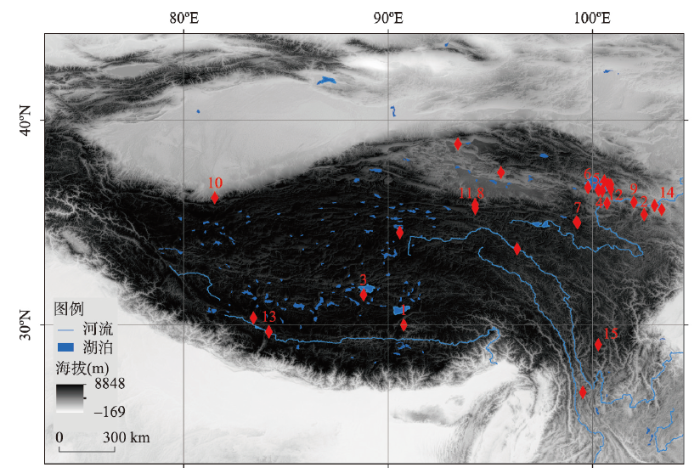
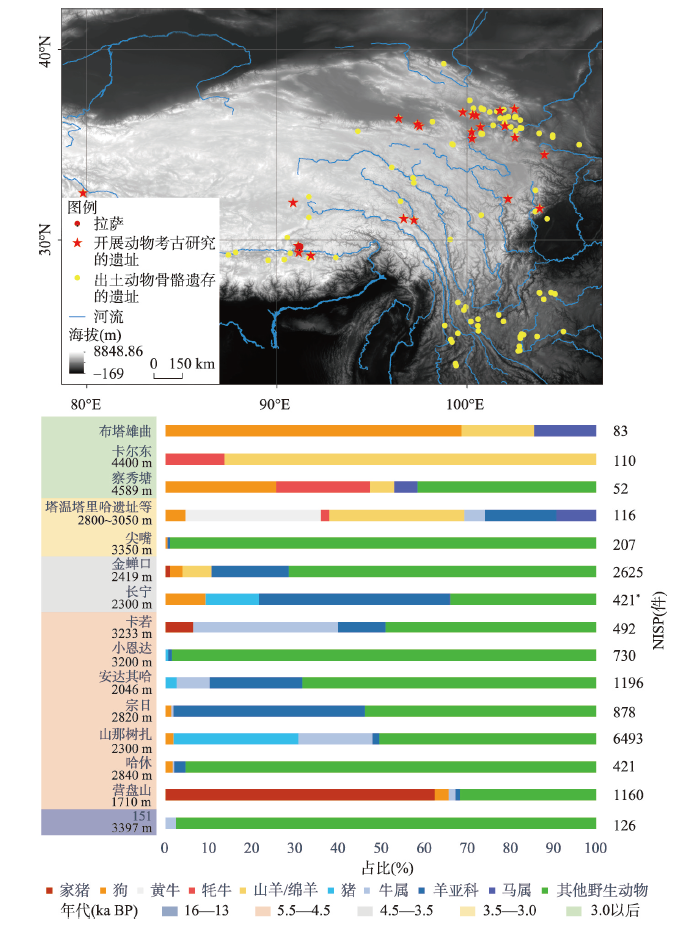
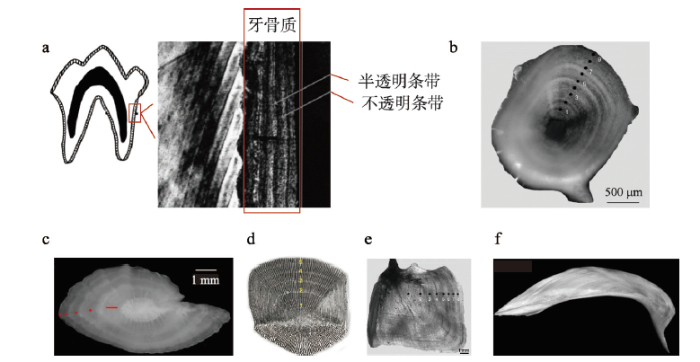


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}