
全新世青藏高原野牦牛适生区模拟及其驯化探讨
|
敖民(1999-), 女, 硕士生, 主要从事环境演变与人类活动研究。E-mail: lm152327@163.com |
收稿日期: 2023-10-16
修回日期: 2024-11-18
网络出版日期: 2025-01-21
基金资助
国家自然科学基金项目(42171165)
青海省科技厅2024年重点研发与转化计划(2024-SF-147)
Simulation of wild yak suitable area and its domestication on the Qinghai-Xizang Plateau in the Holocene
Received date: 2023-10-16
Revised date: 2024-11-18
Online published: 2025-01-21
Supported by
National Natural Science Foundation of China(42171165)
Qinghai Provincial Department of Science and Technology 2024 Key R&D and Transformation Program(2024-SF-147)
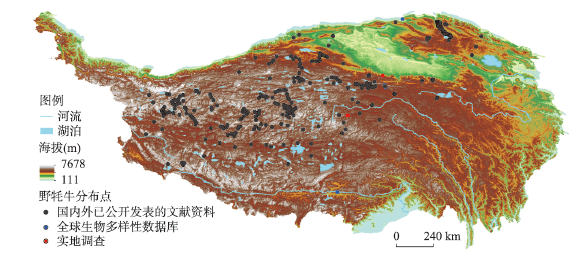
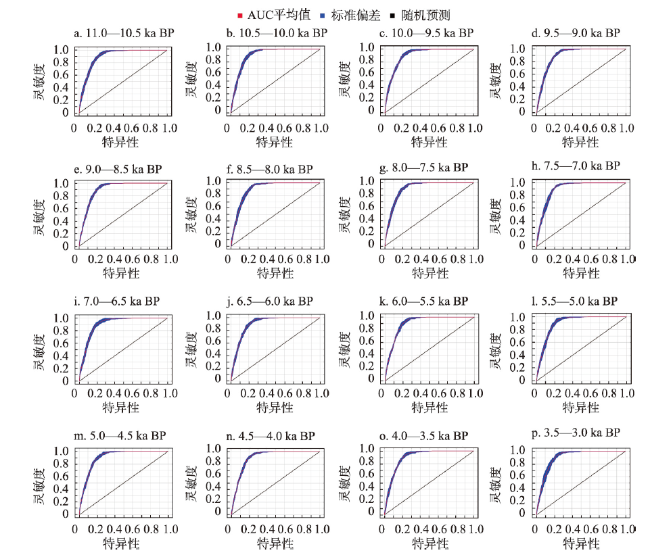
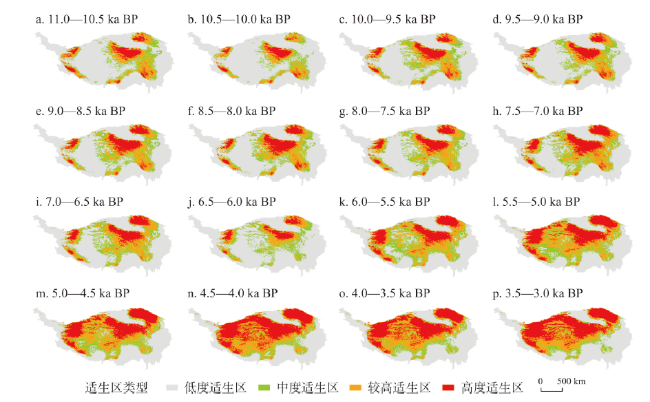
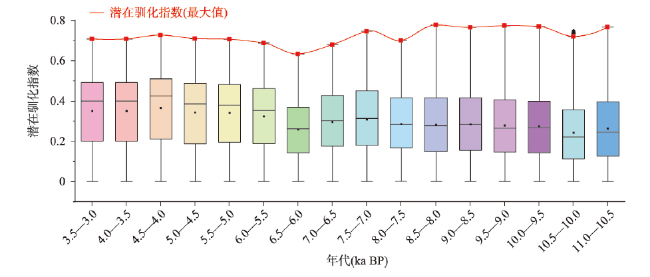
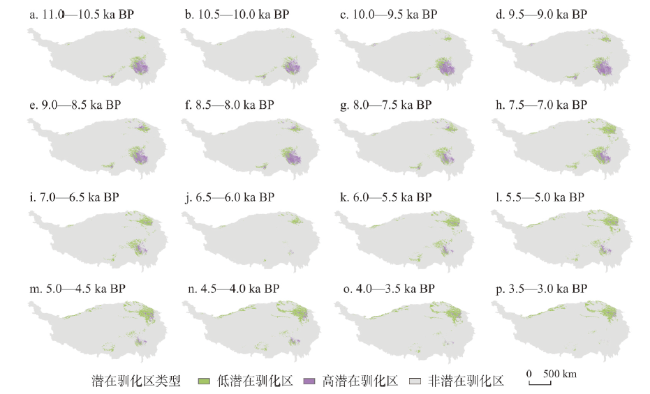
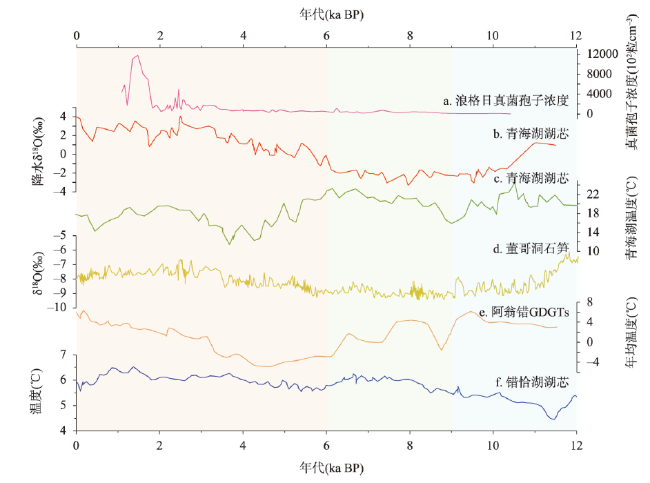
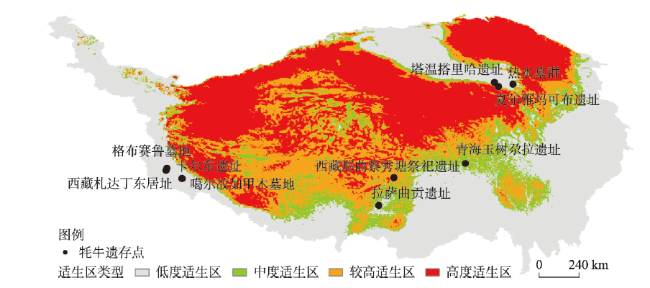
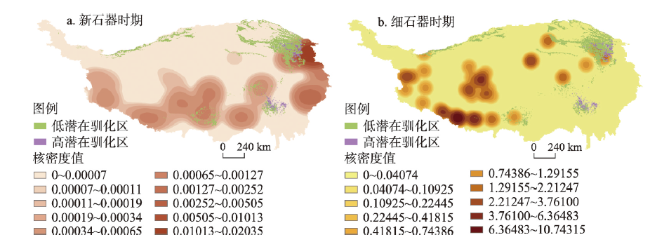
动物的驯化为人类提供了稳定可靠的食物来源,野牦牛是青藏高原的特有物种,在高原地区人类生活中有着不可替代的作用。本文搜集了现今青藏高原野牦牛分布数据291个,利用MaxEnt模型模拟了全新世(11.0—3.0 ka BP)每间隔500 a以来青藏高原野牦牛适生区的时空演变,适生区模拟精度均达到0.9以上。结果表明影响高原上野牦牛适生区的主要环境因子有最冷季度平均温度、降水量变异系数、最湿季度降水量和每年积雪天数。从时空分布来看,全新世早期(11.0—9.0 ka BP)分布面积最小,主要分布在哈拉湖、藏北高原东部、藏南高山谷地东段及昆仑山西段山地高原区等地;全新世早中期(9.0—6.0 ka BP)分布面积介于全新世早期和全新世中晚期之间,主要分布在祁连山脉、藏北高原西部及青南高原等地区。全新世中晚期(6.0—3.0 ka BP)分布面积达到最大值,主要分布在青海湖盆地、藏北高原、羌塘高原、长江—澜沧江上游等地。全新世以来野牦牛的适生区整体呈现逐渐扩大的趋势,其原因可能与气候环境演变有关。结合极端环境指数和野牦牛适生区指数综合计算,推测野牦牛潜在驯化时间在4.5—4.0 ka BP左右,潜在驯化区在青海湖盆地和长江—澜沧江上游及横断山脉。
敖民 , 侯光良 , 陈鸿明 , 文德卓玛 , 金孙梅 , 侯志瑞 , 关佳萌 . 全新世青藏高原野牦牛适生区模拟及其驯化探讨[J]. 地理学报, 2025 , 80(1) : 200 -216 . DOI: 10.11821/dlxb202501013
The domestication of animals has provided humans with a stable food source. The wild yak is a unique species on the Qinghai-Xizang Plateau and plays an irreplaceable role in daily life of the Tibetan people. This study collected 291 localities of wild yaks on the Qinghai-Xizang Plateau and modeled the spatiotemporal evolution of wild yaks on the plateau every 500 years since the Holocene (11.0-3.0 ka BP) by Maxent. The accuracy of the simulation result reached above 0.9. The results indicated that the main environmental factors affecting the suitable habitat of wild yaks on the Qinghai-Xizang Plateau include the average temperature of the coldest quarter, the coefficient variation of precipitation, the precipitation of the wettest quarter, and the accumuation of the annual snowfall. From a spatiotemporal perspective, the distribution area was the smallest in the early Holocene (11.0-9.0 ka BP), mainly distributed in areas like Hala Lake, the eastern part of the northern Tibetan Plateau, the highlands and valleys in the southern Tibetan Plateau, and the highlands in the western Kunlun Mountains. During the early period of the middle Holocene (9.0-6.0 ka BP), the distribution area was located between that of the early and late Holocene, mainly in areas such as the Qilian Mountains, the western part of the northern Tibetan Plateau, and the southern Qinghai Plateau. In the middle to late Holocene (6.0-3.0 ka BP), the distribution area reached its maximum, mainly in areas such as the Qinghai Lake basin, the northern Tibetan Plateau, the Qiangtang Plateau, and the upper reaches of the Yangtze River and Lancang River. In addition, since the Holocene, the habitat of the wild yak has gradually expanded, possibly due to climate and environmental changes. Comprehensive analysis of the Human Living Environment Index and the Wild Yak Habitat Index revealed that the earliest potential domestication time of the wild yak is around 4.5-4.0 ka BP, with potential domestication areas including the Qinghai Lake basin, the upper reaches of the Yangtze River, Lancang River, and the Hengduan Mountains.
表1 环境数据信息表Tab. 1 Environmental data information table |
| 环境数据来源 | Chelsa-Climate数据库 |
|---|---|
| 数据年代范围 | 11.0~3.0 ka BP |
| 空间分辨率 | 1 km(经重采样后分辨率为5 km) |
| 时间分辨率 | 100 a |
| 坐标系 | WGS 84 |
表2 Chelsa-Climate 数据库中的环境变量Tab. 2 Environmental variables in the Chelsa-Climate database |
| 因子代码 | 描述 | 因子代码 | 描述 | 因子代码 | 描述 |
|---|---|---|---|---|---|
| tasmax | 每日最低近地表气温 | bio5 | 最暖月最高温度 | bio14 | 最干月降水量 |
| tasmin | 每日最高近地表气温 | bio6 | 最冷月最高温度 | bio15 | 降水量变异系数 |
| pr | 降水 | bio7 | 年均温变化范围 | bio16 | 最湿季度降水量 |
| scd | 每年积雪天数 | bio8 | 最湿季度平均温度 | bio17 | 最干季度降水量 |
| swe | 雪水当量 | bio9 | 最干季度平均温度 | bio18 | 最暖季度降水量 |
| bio1 | 年平均温度 | bio10 | 最暖季度平均温度 | bio19 | 最冷季度降水量 |
| bio2 | 平均日较差 | bio11 | 最冷季度平均温度 | dem | 海拔 |
| bio3 | 等温性 | bio12 | 年平均降水量 | glz | 冰川高程 |
| bio4 | 温度季节性变化标准 | bio13 | 最湿月降水量 | tz | 空气—温度递减率 |
表3 代入模型的环境变量重要性程度Tab. 3 Importance of the environment variables substituted into the model |
| 变量 | 贡献率(%) | 排列重要性(%) |
|---|---|---|
| Scd(每年积雪天数) | 46.4 | 1.6 |
| bio15(降水量变异系数) | 33.8 | 34.6 |
| bio16(最湿季度降水量) | 15.1 | 15.6 |
| bio7(年均温变化范围) | 3.5 | 9.3 |
| bio11(最冷季度平均温度) | 0.8 | 35.3 |
| bio17(最干季度降水量) | 0.4 | 3.5 |
| [1] |
[张镱锂, 李炳元, 郑度. 论青藏高原范围与面积. 地理研究, 2002, 21(1): 1-8.]
|
| [2] |
[翟欣, 潘志萍, 张春兰, 等. 中国珍稀野生动物手绘图谱. 广州: 中山大学出版社, 2019. 187.]
|
| [3] |
|
| [4] |
[刘艳斌. 野牦牛基因组组装及牦牛泛基因组的构建[D]. 兰州: 兰州大学, 2020.]
|
| [5] |
[赖松家, 王玲, 刘益平, 等. 中国部分牦牛品种线粒体DNA遗传多态性研究. 遗传学报, 2005, 32(5): 463-470.]
|
| [6] |
[郭松长. 牦牛遗传多样性、起源驯化及分类学地位研究: 线粒体DNA D-loop序列的分析[D]. 西宁: 中国科学院西北高原生物研究所, 2006.]
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
[王理中. 牦牛驯化的基因组学证据[D]. 兰州: 兰州大学, 2016.]
|
| [12] |
Institute of Archaeology Chinese Academy of Social Sciences, The Bureau of Cultural Relics Xizang Autonomous Region. Qugong in Lhasa. Beijing: Encyclopedia of China Publishing House, 1999.
[中国社会科学院考古研究所, 西藏自治区文物局. 拉萨曲贡. 北京: 中国大百科全书出版社, 1999.]
|
| [13] |
[屈延华, 雷富民, 尹祚华. 雪雀属鸟类栖息地在中国的分布. 动物学报, 2002, 48(4): 471-479.]
|
| [14] |
[崔绍朋, 罗晓, 李春旺, 等. 基于MaxEnt模型预测白唇鹿的潜在分布区. 生物多样性, 2018, 26(2): 171-176.]
|
| [15] |
[魏临风. 气候变化对青藏高原重点保护动物适宜分布区的影响及其保护状况分析[D]. 金华: 浙江师范大学, 2022.]
|
| [16] |
|
| [17] |
[张镱锂, 李炳元, 刘林山, 等. 再论青藏高原范围. 地理研究, 2021, 40(6): 1543-1553.]
|
| [18] |
[蒋宏伟, 王若勇, 齐纳秀, 等. 野牦牛与家牦牛六个方面比较. 畜牧兽医杂志, 2018, 37(4): 70-71.]
|
| [19] |
[张振西, 林扎西尖措, 高太侦, 等. 基于MaxEnt模型的天峻县大型野生草食动物生境适宜性评价. 中国草食动物科学, 2023, 43(1): 13-18.]
|
| [20] |
[苏旭坤, 董世魁, 刘世梁, 等. 阿尔金山国家级自然保护区基于GIS-生态位模型的野牦牛栖息地辨识研究. 干旱区地理, 2014, 37(6): 1216-1221.]
|
| [21] |
[胡一鸣, 李玮琪, 蒋志刚, 等. 羌塘、可可西里无人区野牦牛种群数量和分布现状. 生物多样性, 2018, 26(2): 185-190.]
|
| [22] |
|
| [23] |
|
| [24] |
[刘红彩, 赵纳勋, 庄钰琪, 等. 基于MaxEnt模型的秦岭山地斑羚生境适宜性评价. 生态学报, 2022, 42(10): 4181-4188.]
|
| [25] |
|
| [26] |
[殷颖璇, 徐安远, 潘筱雯, 等. 基于优化MaxEnt模型预测玫瑰蜗牛(Euglandina Rosea)在中国的潜在分布区. 中国媒介生物学及控制杂志, 2023, 34(2): 137-144.]
|
| [27] |
|
| [28] |
[金孙梅, 侯光良, 陈晓良, 等. 青藏高原末次冰消期以来人类活动的时空演化特征及其原因探讨. 第四纪研究, 2022, 42(1): 223-235.]
|
| [29] |
[卢松, 张小军, 张业臣. 徽州传统村落的时空分布及其影响因素. 地理科学, 2018, 38(10): 1690-1698.]
|
| [30] |
[孔维尧, 李欣海, 邹红菲. 最大熵模型在物种分布预测中的优化. 应用生态学报, 2019, 30(6): 2116-2128.]
|
| [31] |
[王运生, 谢丙炎, 万方浩, 等. ROC曲线分析在评价入侵物种分布模型中的应用. 生物多样性, 2007, 15(4): 365-372.]
|
| [32] |
[宋慧芳, 党晓宏, 高永, 等. 基于MaxEnt模型的内蒙古自治区樟子松潜在分布研究. 四川农业大学学报, 2023, 41(2): 203-208.]
|
| [33] |
|
| [34] |
[刘务林, 乔治⋅B⋅夏勒. 野牦牛的分布和现状. 西藏科技, 2003(11): 17-23.]
|
| [35] |
[侯光良, 鄂崇毅, 肖景义. 青藏高原全新世降水序列的集成重建. 地理科学进展, 2012, 31(9): 1117-1123.]
|
| [36] |
|
| [37] |
[盛淑蓉, 魏海成, 山发寿, 等. 哈拉湖流域表土花粉与植被关系研究. 盐湖研究, 2014, 22(2): 20-25.]
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
[魏海成, 鄂崇毅, 段荣蕾, 等. 真菌孢子记录的全新世中期以来青藏高原东北部地区畜牧活动历史. 中国科学: 地球科学, 2021, 51(11): 1907-1922.]
|
| [44] |
[刘兴起, 沈吉, 王苏民, 等. 青海湖16ka以来的花粉记录及其古气候古环境演化. 科学通报, 2002, 47(17): 1351-1355.]
|
| [45] |
[陈晓良. 长江—澜沧江源区史前人类活动与环境适应研究[D]. 西宁: 青海师范大学, 2022.]
|
| [46] |
[李凡, 侯光良, 鄂崇毅, 等. 青藏高原全新世气温序列的集成重建. 干旱区研究, 2015, 32(4): 716-725.]
|
| [47] |
|
| [48] |
[李拓宇, 任小林, 廖奕楠, 等. 雅鲁藏布江中游及昌果沟遗址古环境分析. 第四纪研究, 2020, 40(2): 547-555.]
|
| [49] |
|
| [50] |
[韩建华, 白文龙, 甄强, 等. 青海都兰县热水墓群2018血渭一号墓. 考古, 2021, 67(8): 45-70.]
|
| [51] |
|
| [52] |
[吕红亮. 西藏阿里地区丁东居住遗址发掘简报. 考古, 2007, 53(11): 36-46.]
|
| [53] |
|
| [54] |
Henan Provincial Institute of Cultural Heritage and Archaeology. Zooarchaeology (Vol. 3). Beijing: Cultural Relics Publishing House, 2019.
[河南省文物考古研究院.动物考古(第3辑). 北京: 文物出版社, 2019.]
|
| [55] |
[仝涛, 李林辉, 赤列次仁, 等. 西藏阿里地区故如甲木墓地和曲踏墓地. 考古, 2015, 61(7): 29-50.]
|
| [56] |
[张正为, 吕红亮. 西藏西部阿里卡尔东遗址2013年试掘出土动物遗存鉴定与分析. 藏学学刊, 2017, 13(1): 252-272.]
|
| [57] |
[胡松梅, 张建林. 西藏那曲察秀塘祭祀遗址哺乳动物遗存及其意义. 2007年中国郑州动物考古国际学术研讨会论文集. 郑州: 河南省文物局, 2007: 241-251.]
|
| [58] |
[陈顺港, 高玉, 陈宁博, 等. 青藏高原牦牛驯化的考古学与遗传学研究进展及展望. 科学通报, 2024, 69(11): 1417-1428.]
|
| [59] |
[吴汝祥. 青海都兰县诺木洪搭里他里哈遗址调查与试掘. 考古学报, 1963, 28(1): 17-44.]
|
| [60] |
[李永宪. 西藏原始艺术. 石家庄: 河北教育出版社, 2000: 179-180.]
|
| [61] |
[王永军. 玉树通天河流域岩画中牦牛图像研究. 中国藏学, 2018, 31(1): 77-88.]
|
| [62] |
[魏海成. 青海湖盆地畜牧活动起源与发展历史探究: 以江西沟2号遗址为例. 盐湖研究, 2020, 28(4): 10-21.]
|
| [63] |
|
| [64] |
[金孙梅. 通天河流域史前人类活动及生存策略研究[D]. 西宁: 青海师范大学, 2024.]
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
[王建, 夏欢, 姚娟婷, 等. 青藏高原末次冰消期狩猎采集人群的生存策略研究. 中国科学: 地球科学, 2020, 50(3): 380-390.]
|
| [70] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}