
30 ka BP以来大兴安岭漠河盆地多年冻土区气候与环境变化
|
刘锐(1989-), 男, 黑龙江哈尔滨人, 博士生, 主要从事寒区第四纪环境演变研究。E-mail: 7414369@qq.com |
收稿日期: 2023-12-01
修回日期: 2024-08-19
网络出版日期: 2024-09-27
基金资助
科技基础资源调查专项项目(2022FY100701)
国家自然科学基金联合基金重点项目(U20A2082)
国家自然科学基金项目(41971151)
哈尔滨师范大学博士研究生创新基金项目(HSDBSCX2022-07)
Climate and environmental changes in the Mohe Basin permafrost region of the Greater Hinggan Mountains since 30 ka BP
Received date: 2023-12-01
Revised date: 2024-08-19
Online published: 2024-09-27
Supported by
Science & Technology Fundamental Resources Investigation Program(2022FY100701)
Key Joint Program of National Natural Science Foundation of China(U20A2082)
National Natural Science Foundation of China(41971151)
Doctoral Innovation Foundation of Harbin Normal University(HSDBSCX2022-07)
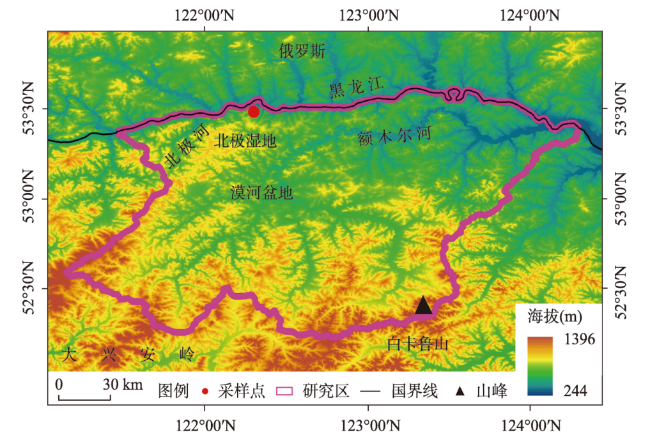
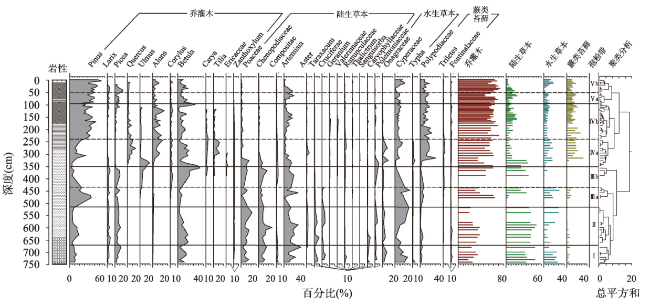
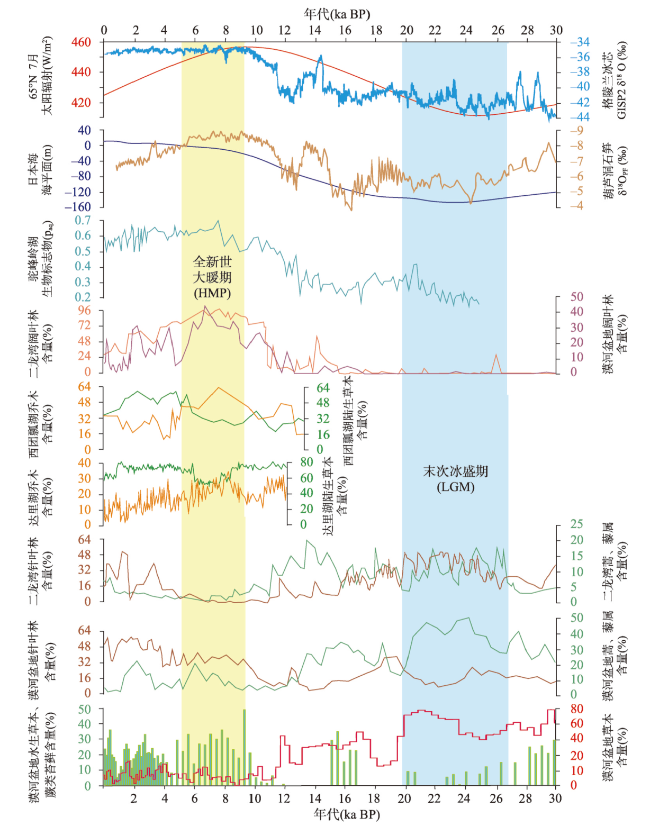
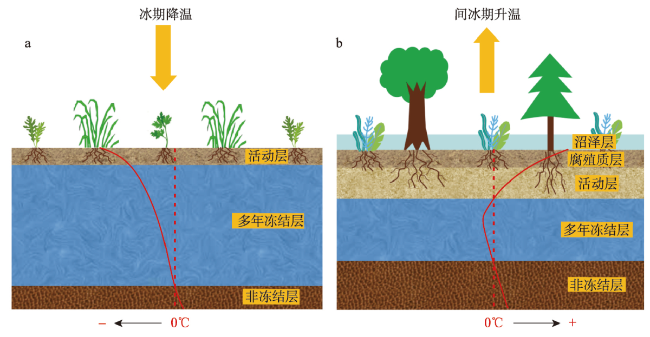
多年冻土对全球变化非常敏感,利用植被演替揭示多年冻土区气候与环境变化具有重要意义。本文以大兴安岭漠河盆地多年冻土岩芯孢粉学为证据,通过AMS14C构建了年代学框架,重建多年冻土区更新世晚期以来植被演替与气候历史,分析了植被演替对多年冻土区环境变化的响应。结果显示孢粉组合较好地反映漠河盆地和外围山地植被组成格局,依据区域关键植被对气候指示关系,漠河盆地30 ka BP以来植被演替指示气候发生了5次显著变化过程:30.0—27.1 ka BP形成草甸湿地景观,指示气候寒冷湿润;27.1—20.5 ka BP形成针叶林草原景观,指示气候相对寒冷干旱;20.5—11.3 ka BP由针叶林湿地向针阔混交林湿地景观转变,指示气温开始转暖,但气候仍寒冷湿润;11.3—1.9 ka BP形成针阔混交林湿地景观,指示气候温暖湿润;1.9 ka BP至今形成针叶林湿地景观,指示气候冷凉湿润。通过对比分析发现,影响植被演替的因素不仅包括纬度和海陆位置上的气候差异,还有多年冻土本身引起的区域环境效应,植被演替对冰期、间冰期多年冻土变化具有不同响应机制。在末次冰盛期,由于多年冻土冻结和扩张,导致多年冻土环境更能适应耐寒耐旱、根系较浅的蒿属、藜属等陆生草本繁殖,干旱草原扩张;在全新世大暖期,多年冻土的融化和退缩不仅为植被提供了良好的生境,还有利于湿地的潜育化和面积扩大,促进水生植被、蕨类苔藓和乔木的大量繁殖,形成森林湿地景观。因此,由于多年冻土的存在,放大了气候的水热特性,植被对多年冻土区环境变化的响应更为敏感。
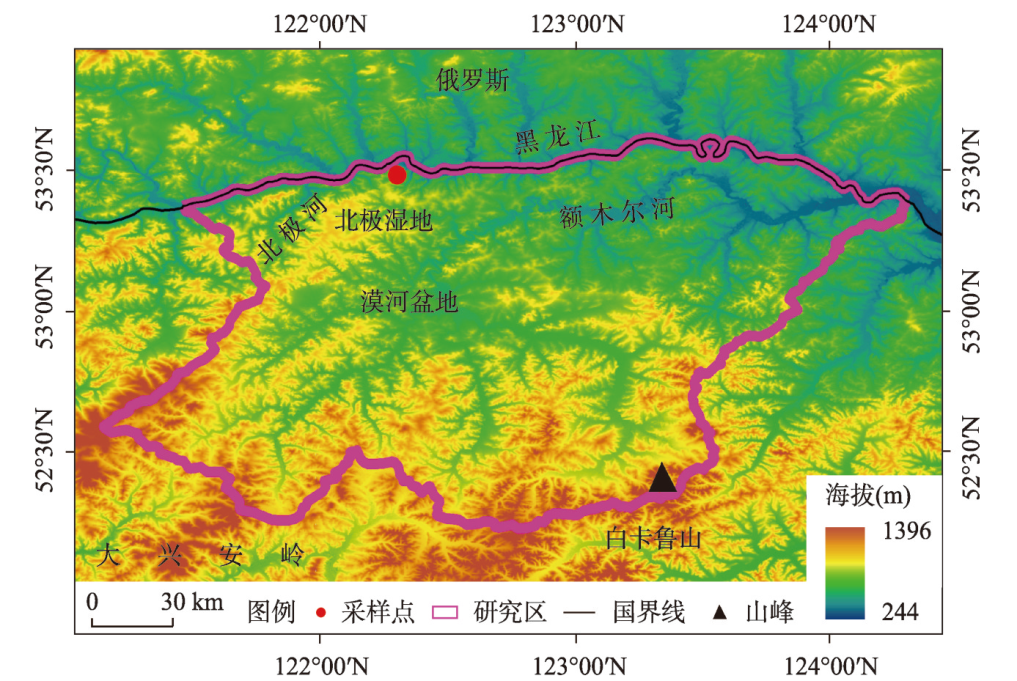
刘锐 , 臧淑英 , 赵林 , 吴晓东 , 刘立新 , 吴少强 , 李天瑞 , 张紫豪 , 何俭翔 , 李彪 , 张博雄 , 程小峰 . 30 ka BP以来大兴安岭漠河盆地多年冻土区气候与环境变化[J]. 地理学报, 2024 , 79(9) : 2280 -2296 . DOI: 10.11821/dlxb202409008
Permafrost is highly sensitive to global changes, and it is important to reveal climate and environmental changes in permafrost regions using vegetation succession. Based on palynological evidence and AMS14C dating from a sediment core, vegetation succession and climate history in the Late Pleistocene of the Mohe Basin (MHB) in the Greater Hinggan Mountain permafrost region was reconstructed. The response of vegetation to environmental changes was further investigated. Results showed that pollen assemblages effectively reflect the vegetation composition in the MHB and surrounding mountainous areas. Based on the relationship between key regional vegetation and climate indicators, vegetation has undergone significant historical changes: from 30.0 to 27.1 ka BP, a meadow wetland landscape showed a cold and humid climate; from 27.1 to 20.5 ka BP, a coniferous forest-grassland landscape indicated a relatively cold and arid climate; from 20.5 to 11.3 ka BP, there was a transition from coniferous forest wetlands to mixed coniferous-broadleaved forest wetlands, indicating the beginning of a warming although the climate remained cold and humid; from 11.3 to 1.9 ka BP, a mixed coniferous-broadleaved forest wetland landscape demonstrated a warm and humid climate; from 1.9 ka BP to the present, a coniferous forest wetland landscape revealed a cool and humid climate. A comparative analysis showed that factors influencing vegetation succession include not only climate conditions related to latitude and land-sea location but also regional factors caused by permafrost itself. Vegetation succession exhibited different response mechanisms to permafrost changes during glacial and interglacial periods. During the Last Glacial Maximum, permafrost expansion benefited the growth of cold and drought-resistant terrestrial herbs with shallow roots such as Artemisia and Chenopodiaceae, leading to the expansion of arid grasslands. During the Holocene Megathermal Period, permafrost degradation provided favorable habitats for vegetation, and also facilitated the formation and expansion of wetlands, promoting the extensive proliferation of aquatic plants, ferns, mosses, and trees, forming a forest wetland landscape. Therefore, permafrost presence enhances the effects of water and heat under different climate conditions on vegetation, making vegetation more sensitive to environmental changes in permafrost regions.
Key words: Mohe Basin; Late Pleistocene; permafrost; vegetation response; paleoclimate
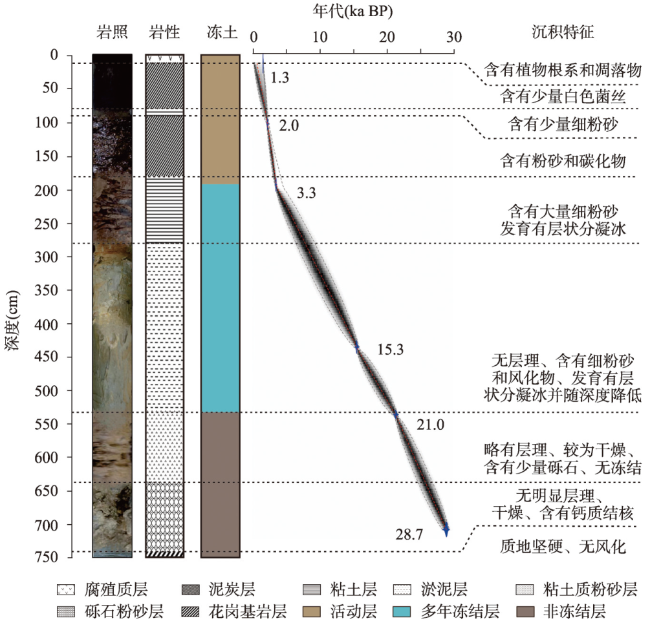
表1 漠河盆地BJC-3岩芯AMS14C年龄Tab. 1 AMS14C dating of BJC-3 core in the Mohe Basin |
| 取样编号 | 测试编号 | 取样冻土层位 | 测样材料 | 深度 (cm) | 14C表观年龄 (ka BP) | 14C校正年龄 (2σ ka BP) | 14C中值年龄 (ka BP) |
|---|---|---|---|---|---|---|---|
| XAB-1 | 621829 | 活动层 | 泥炭 | 20 | 1.4 | 1.2—1.3 | 1.3 |
| XAB-2 | 621831 | 活动层 | 泥炭 | 110 | 2.0 | 1.9—2.1 | 2.0 |
| XAB-3 | 621832 | 活动层 | 有机物 | 190 | 3.1 | 3.2—3.3 | 3.3 |
| XAB-4 | 621833 | 多年冻结层 | 淤泥 | 430 | 12.8 | 15.1—15.5 | 15.3 |
| XAB-5 | 621835 | 多年冻结层 | 淤泥 | 530 | 17.4 | 20.8—21.2 | 21.0 |
| XAB-6 | 621834 | 非冻结层 | 有机物 | 710 | 24.5 | 28.6—28.9 | 28.7 |
注:14C半衰期为5568年,起始年龄自1950年算起。 |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
[邱国庆, 程国栋. 中国的多年冻土: 过去与现在. 第四纪研究, 1995, 15(1): 13-22.]
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
[童国榜, 羊向东, 王苏民, 等. 扎赍诺尔晚第四纪孢粉植物群的生态类型探讨. 地理学报, 1997, 52(1): 72-79.]
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
[常晓丽, 金会军, 王永平, 等. 植被对多年冻土的影响研究进展. 生态学报, 2012, 32(24): 7981-7990.]
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
[伍婧, 刘强. 晚冰期以来月亮湖孢粉记录反映的古植被与古气候演化. 地球科学, 2012, 37(5): 947-954.]
|
| [18] |
[王利贤, 刘强, 游海涛. 晚冰期以来月亮湖生物硅含量变化的古气候意义. 地球与环境, 2012, 40(3): 328-335.]
|
| [19] |
[伍婧, 刘强. 晚冰期月亮湖炭屑记录反映的古气候演化. 地质科学, 2013, 48(3): 860-869.]
|
| [20] |
[刘嘉丽, 刘强, 储国强, 等. 大兴安岭四方山天池15.4 ka B.P. 以来湖泊沉积记录. 第四纪研究, 2015, 35(4): 901-912.]
|
| [21] |
[刘嘉丽, 刘强, 伍婧, 等. 大兴安岭四方山天池全新世以来沉积物正构烷烃分布、单体碳同位素特征及古环境意义. 湖泊科学, 2017, 29(2): 498-511.]
|
| [22] |
|
| [23] |
[李梦真, 介冬梅, 李楠楠, 等. 中全新世以来大兴安岭中段红水泡泥炭地硅藻组合记录的古环境演化. 微体古生物学报, 2019, 36(2): 163-174.]
|
| [24] |
[杨月, 介冬梅, 李楠楠, 等. 大兴安岭红水泡泥炭地腐殖化度与总有机碳记录的5 ka BP以来的气候变化. 湿地科学, 2019, 17(4): 424-435.]
|
| [25] |
[李雪林. 内蒙古阿尔山天池小冰期以来湖泊钻孔孢粉特征及植被演替[D]. 北京: 中国地质大学, 2012.]
|
| [26] |
[崔巧玉, 赵艳. 大兴安岭阿尔山天池湖泊沉积物记录的全新世气候突变. 第四纪研究, 2019, 39(6): 1346-1356.]
|
| [27] |
[冷程程, 赵成, 崔巧玉, 等. 大兴安岭阿尔山天池沉积物中正构烷烃记录揭示的全新世古气候变化. 第四纪研究, 2021, 41(4): 976-985.]
|
| [28] |
|
| [29] |
|
| [30] |
[马雪云, 魏志福, 王永莉, 等. 末次冰盛期以来东北地区霍拉盆地湖泊沉积物记录的C3/C4植被演化. 第四纪研究, 2018, 38(5): 1193-1202.]
|
| [31] |
|
| [32] |
|
| [33] |
[李宜垠, 李博闻, 徐鑫. 大兴安岭北部满归泥炭孢粉重建的过去2100年古气候. 第四纪研究, 2019, 39(4): 1034-1041.]
|
| [34] |
[王丽丽, 邵雪梅, 黄磊, 等. 黑龙江漠河兴安落叶松与樟子松树轮生长特性及其对气候的响应. 植物生态学报, 2005, 29(3): 280-285.]
|
| [35] |
[刘畅往, 裴发根, 仇根根, 等. 东北漠河盆地北部冻土分布特征. 物探与化探, 2017, 41(6): 1204-1214.]
|
| [36] |
[胡理乐, 李俊生, 刘文慧, 等. 大兴安岭常见植物彩色图鉴. 北京: 科学出版社, 2014.]
|
| [37] |
|
| [38] |
[王伏雄, 钱南芬, 张玉龙, 等. 中国植物花粉形态, 2版. 北京: 科学出版社, 1995.]
|
| [39] |
[唐领余, 毛礼米, 舒军武, 等. 中国第四纪孢粉图鉴. 北京: 科学出版社, 2016.]
|
| [40] |
|
| [41] |
|
| [42] |
[李良, 孙丰月, 李碧乐, 等. 漠河盆地二十二站组砂岩形成时代及物源区构造环境判别. 地球科学, 2017, 42(1): 35-52.]
|
| [43] |
|
| [44] |
|
| [45] |
[孙湘君, 王琫瑜, 宋长青. 中国北方部分科属花粉—气候响应面分析. 中国科学: 地球科学, 1996, 26(5): 431-436.]
|
| [46] |
|
| [47] |
|
| [48] |
[周以良. 中国东北植被地理. 北京: 科学出版社, 1997: 49-51.]
|
| [49] |
[周以良. 中国大兴安岭植被. 北京: 科学出版社, 1991: 88-91.]
|
| [50] |
[沈才明, 唐领余. 全新世长白山和小兴安岭气候:中国全新世大暖期气候与环境. 北京: 中国海洋出版社, 1992.]
|
| [51] |
|
| [52] |
|
| [53] |
[崔巧玉, 赵艳, 秦锋, 等. 中国东北表土花粉组合特征及其在古气候重建中的意义. 中国科学: 地球科学, 2020, 50(1): 134-148.]
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
[李小强, 赵超, 周新郢. 末次盛冰期以来中国东北地区特征时期植被格局. 中国科学: 地球科学, 2019, 49(8): 1213-1230.]
|
| [61] |
[李楠楠. 中国东北龙岗地区新仙女木事件以来植被动态对气候变化的响应[D]. 长春: 东北师范大学, 2020.]
|
| [62] |
|
| [63] |
|
| [64] |
[马瑞丰, 张威, 金培红, 等. 辽宁南部金州地区距今13.5 ka以来古植被古气候变化. 第四纪研究, 2021, 41(1): 43-50.]
|
| [65] |
[王娜, 许清海, 张生瑞, 等. 白洋淀地区晚冰期以来的气候和环境演变. 地理学报, 2022, 77(5): 1195-1210.]
|
| [66] |
|
| [67] |
[邢伟, 鲍锟山, 韩冬雪, 等. 全新世以来东北地区沼泽湿地发育过程及其对气候变化的响应. 湖泊科学, 2019, 31(5): 1391-1402.]
|
| [68] |
|
| [69] |
[施雅风, 孔昭宸, 王苏民, 等. 中国全新世大暖期鼎盛阶段的气候与环境. 中国科学: B辑, 1993, 23(8): 865-873.]
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
[刘玉英. 晚更新世晚期以来二龙湾玛珥湖植被与环境演化研究[D]. 长春: 吉林大学, 2009.]
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
[金会军, 金晓颖, 何瑞霞, 等. 两万年来的中国多年冻土形成演化. 中国科学: 地球科学, 2019, 49(8): 1197-1212.]
|
| [79] |
|
| [80] |
|
| [81] |
[孙广友. 试论沼泽与冻土的共生机理: 以中国大小兴安岭地区为例. 冰川冻土, 2000, 22(4): 309-316.]
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}