
近200年青藏高原东南部真布错硅藻记录与环境变化
|
何晋娜(1987-), 女, 山西运城人, 博士, 主要从事湖泊生态研究。E-mail: hejn0201@163.com |
收稿日期: 2023-06-30
修回日期: 2024-03-15
网络出版日期: 2024-05-31
基金资助
中国科学院基础前沿科学研究计划从0到1原始创新项目(ZDBS-LY-DQC006)
国家自然科学基金项目(42071115)
国家自然科学基金项目(41877460)
国家自然科学基金项目(41871070)
第二次青藏高原综合科学考察研究(2019QZKK0601)
西藏自治区自然科学基金项目重点项目(XZ202301ZR0014G)
Diatom records and environmental changes over the past 200 years at Zhenbu Co Lake, southeastern Tibetan Plateau
Received date: 2023-06-30
Revised date: 2024-03-15
Online published: 2024-05-31
Supported by
Basic Frontier Science Research Program From 0 to 1 Original Innovation Project of Chinese Academy of Sciences(ZDBS-LY-DQC006)
National Natural Science Foundation of China(42071115)
National Natural Science Foundation of China(41877460)
National Natural Science Foundation of China(41871070)
The Second Tibetan Plateau Scientific Expedition and Research Program(2019QZKK0601)
Natural Science Foundation Key Project of Tibet Autonomous Region(XZ202301ZR0014G)
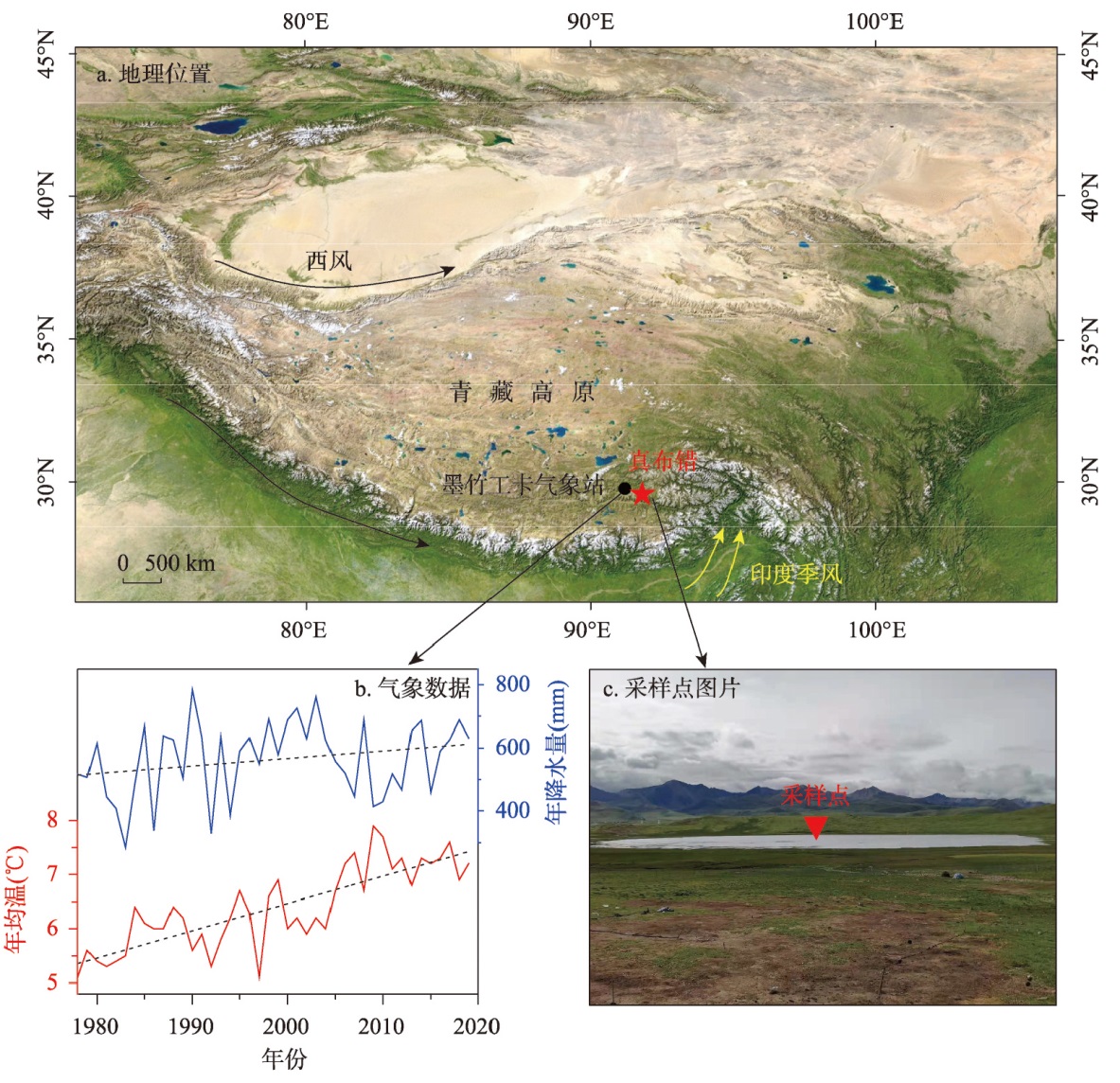
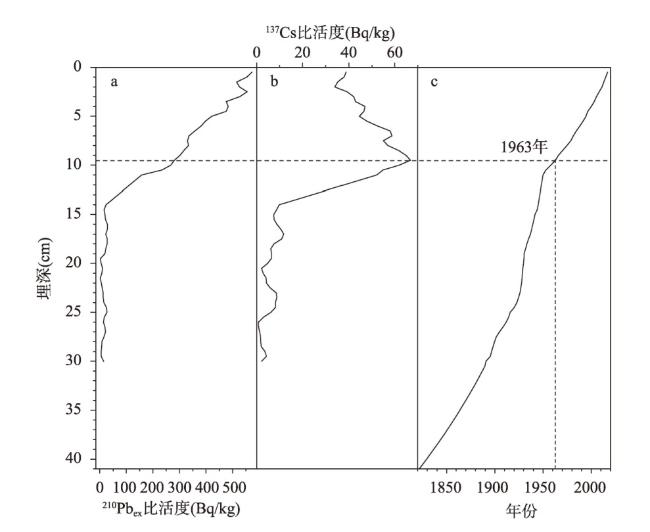
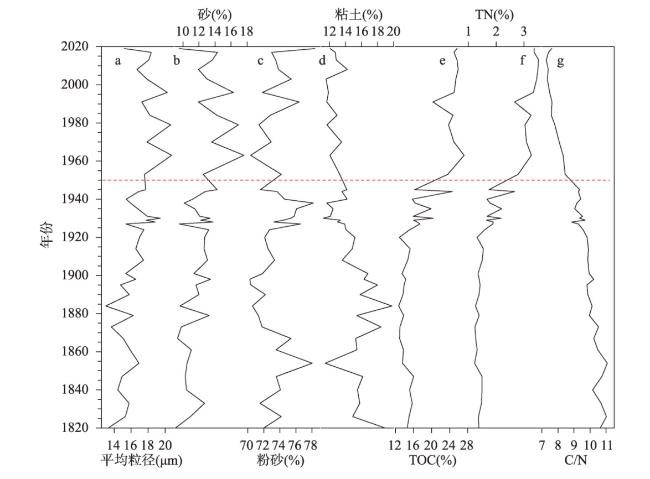
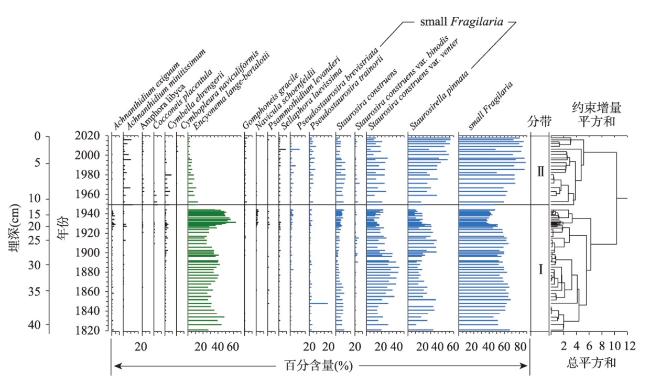
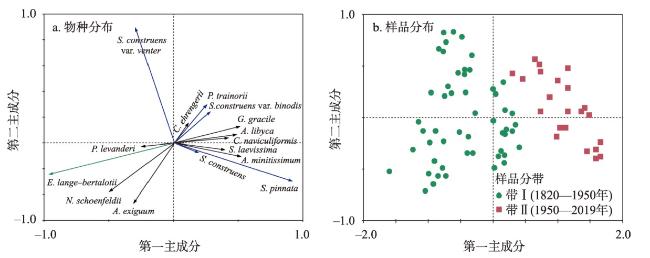
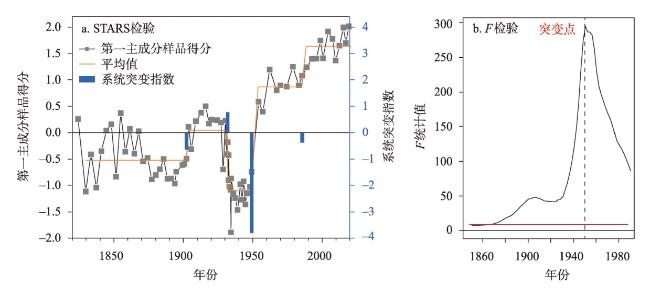
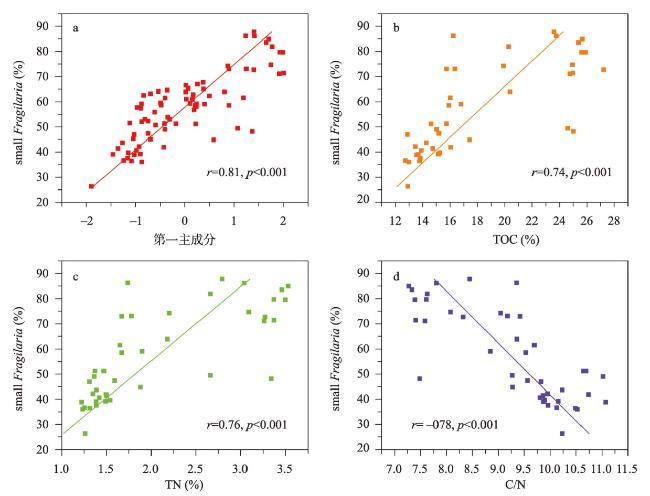
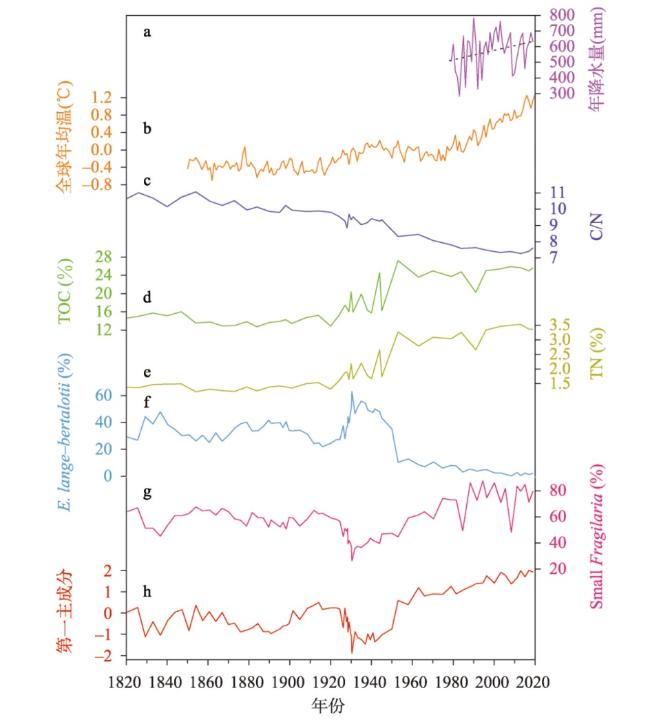
近200年来青藏高原湖泊生态系统对气候变暖的响应研究多集中在深水湖泊,而对大量浅水湖泊生态系统的研究较为匮乏。本文以青藏高原东南部浅水湖真布错为研究对象,通过对沉积物柱芯的210Pb、137Cs测年和高分辨率硅藻鉴定分析,重建该湖过去200年来硅藻属种组合变化,并结合沉积物总有机碳(TOC)含量、总氮(TN)含量、碳氮比(C/N)等指标,探讨湖泊生态系统对气候变化等因素的响应。结果显示,1820—1950年硅藻组成中水生植物附生种Encyonema lange-bertalotii的百分含量较高,表明真布错水生植物较繁盛,湖水透明度高。1950年以来,硅藻组合发生了突变,附生种Encyonema lange-bertalotii的百分含量快速下降,耐低光照的small Fragilaria成为优势属,表明水生植物衰退,湖水透明度降低;同时TOC和TN含量升高,C/N减小,指示湖泊生产力和营养水平升高,藻类在湖泊有机质中占比增加。气温快速升高、降水量增加(冲刷流域更多营养物质进入湖泊)和人类活动增强(流域内出现新建寺庙)可能共同导致湖泊生产力升高,藻类繁盛,水生植物衰退,水体透明度下降,最终引起硅藻群落的突变。因此,在青藏高原气候暖湿化背景下,浅水湖泊可能面临生态系统的转变,未来应该加强该类湖泊的监测与保护。本文记录的高原浅水湖泊生态系统突变时间与中国和全球其他地区基本一致,这也为确定人类世的地层标记提供了新的证据。青藏高原湖泊可以作为研究人类世的天然实验室。
何晋娜 , 索娜卓嘎 , 张继峰 , 曹鹏熙 , 王俊伟 , 袁艳丽 , 葛梦娟 , 杨凤琴 , 刘建宝 . 近200年青藏高原东南部真布错硅藻记录与环境变化[J]. 地理学报, 2024 , 79(5) : 1231 -1245 . DOI: 10.11821/dlxb202405009
Currently, most studies on the response of lake ecosystems on the Tibetan Plateau to climate warming over the past 200 years have focused on deep lakes, whereas studies on ecosystems characterized by many shallow lakes are scarce. In this paper, we selected the shallow lake Zhenbu Co on the southeastern Tibetan Plateau as the research object. Based on 210Pb and 137Cs dating of sediments and high-resolution diatom identification, the changes in the diatom assemblage over the past 200 years were reconstructed. Combined with total organic carbon (TOC), total nitrogen (TN) and carbon-nitrogen ratio (C/N) of the sediments, the response of the lake ecosystem to climate change was explored. The results showed that during 1820-1950 AD, the content of Encyonema lange-bertalotii in sediment was high, indicating the abundant aquatic plants and high lake-water transparency in Zhenbu Co. Since 1950 AD, the diatom assemblages shifted, decreasing rapidly in Encyonema lange-bertalotii content and increasing in small Fragilaria content, which indicated a decline of aquatic plants and decreased lake-water transparency. Meanwhile, the contents of TOC and TN increased, and the C/N decreased, which indicated that the productivity and nutrient level of the lake increased, and the percentage of algae in the lake organic matter increased. Rapidly rising temperature, increased precipitation (i.e., more nutrients entering into the lake), and enhanced human activities (i.e., the appearance of new temples in the watershed) may have jointly led to higher lake productivity and algal bloom content, as well as lower aquatic plant content and decreased water clarity. Eventually, the diatom assemblages abruptly shifted. Based on our results, shallow lakes may face ecosystem shifts in the context of climate warming and humidification on the Tibetan Plateau, and the monitoring and protection of such lakes should be strengthened in the future. The timeline of the abrupt shift of the plateau shallow lake ecosystem in this paper is basically consistent with that in China and other parts of the world, which also provides new evidence for determining the stratigraphic marker of the Anthropocene. Thus, the lakes of the Tibetan Plateau can serve as natural laboratories for studying the Anthropocene.
Key words: Tibetan Plateau; shallow lake; diatom; paleoecology; climate change
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
[秦伯强, 高光, 朱广伟, 等. 湖泊富营养化及其生态系统响应. 科学通报, 2013, 58(10): 855-864.]
|
| [6] |
|
| [7] |
|
| [8] |
[孙鸿烈, 郑度, 姚檀栋, 等. 青藏高原国家生态安全屏障保护与建设. 地理学报, 2012, 67(1): 3-12.]
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
[张国庆. 青藏高原湖泊变化遥感监测及其对气候变化的响应研究进展. 地理科学进展, 2018, 37(2): 214-223.]
|
| [16] |
[张鑫, 吴艳红, 张鑫. 1972—2012年青藏高原中南部内陆湖泊的水位变化. 地理学报, 2014, 69(7): 993-1001.]
|
| [17] |
[姚晓军, 李龙, 赵军, 等. 近10年来可可西里地区主要湖泊冰情时空变化. 地理学报, 2015, 70(7): 1114-1124.]
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
[姚檀栋, 朴世龙, 沈妙根, 等. 印度季风与西风相互作用在现代青藏高原产生连锁式环境效应. 中国科学院院刊, 2017, 32(9): 976-984.]
|
| [25] |
[
|
| [26] |
[
|
| [27] |
[
|
| [28] |
[
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
[刘妍, 王全喜, 施之新. 大兴安岭达尔滨湖桥弯藻科(硅藻门)中国新记录植物. 武汉植物学研究, 2007, 25(6): 565-571.]
|
| [48] |
|
| [49] |
[胡竹君. 青藏高原东南缘高山树线湖泊生态变化与驱动机制[D]. 南京: 中国科学院大学南京湖泊与地理研究所, 2013.]
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
[陈敬安, 万国江, 汪福顺, 等. 湖泊现代沉积物碳环境记录研究. 中国科学: 地球科学, 2002, 32(1): 73-80.]
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
[顾峰雪, 黄玫, 张远东, 等. 1961—2010年中国区域氮沉降时空格局模拟研究. 生态学报, 2016, 36(12): 3591-3600.]
|
| [75] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}