
鄱阳湖草洲景观连通性对植物多样性的影响
|
张成(1991-), 男, 安徽合肥人, 博士生, 主要从事土地利用与景观生态研究。E-mail: zhangchengstudy@126.com |
收稿日期: 2023-05-19
修回日期: 2023-11-01
网络出版日期: 2024-01-29
基金资助
国家自然科学基金项目(42261021)
国家自然科学基金项目(41961036)
Effects of grassland connectivity on plant diversity in Poyang Lake
Received date: 2023-05-19
Revised date: 2023-11-01
Online published: 2024-01-29
Supported by
National Natural Science Foundation of China(42261021)
National Natural Science Foundation of China(41961036)
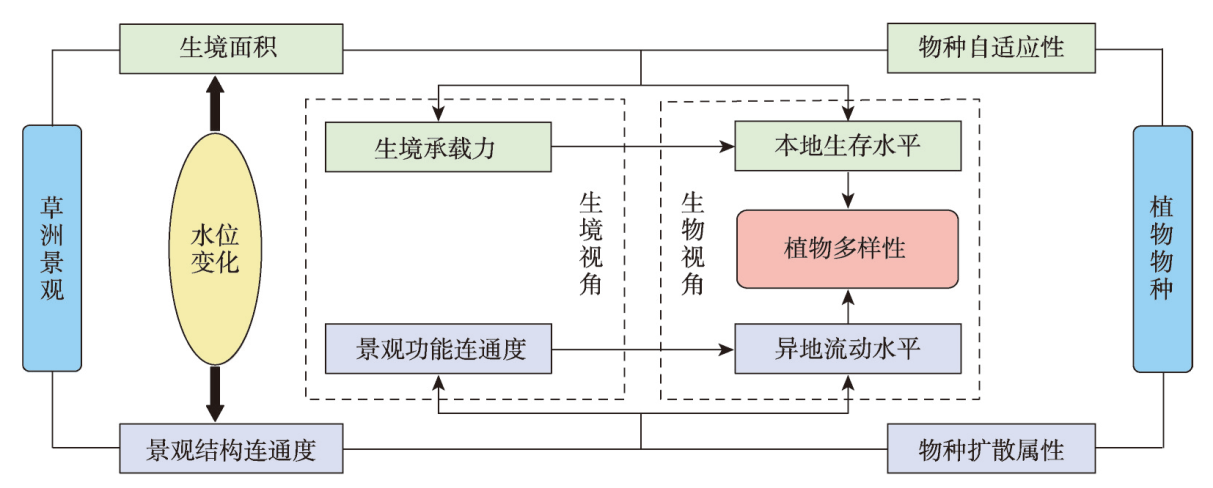
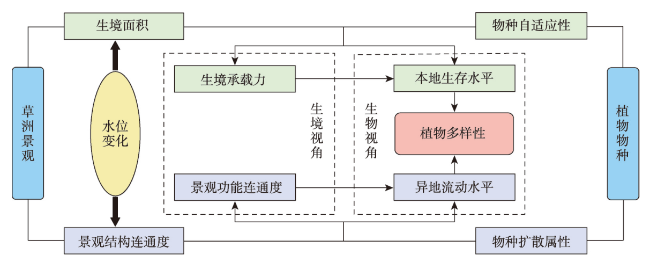
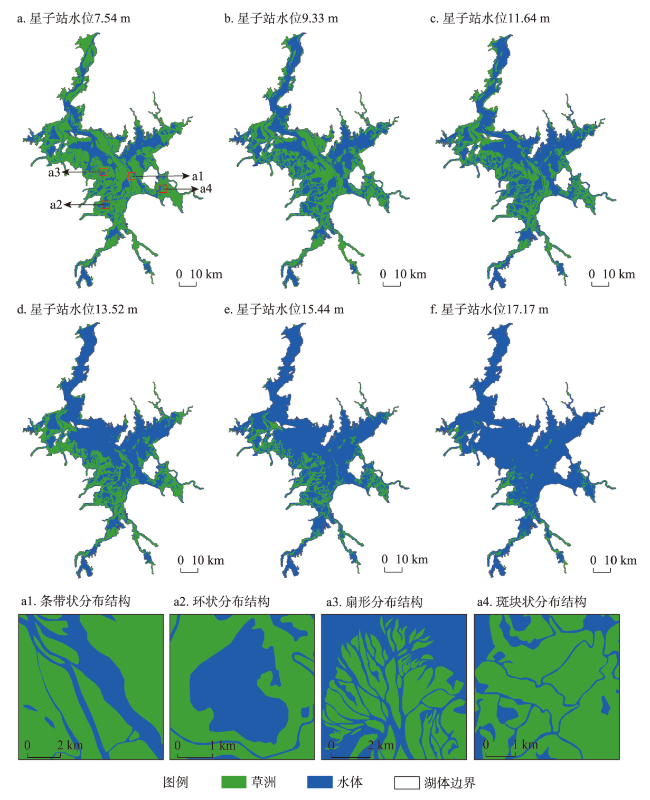
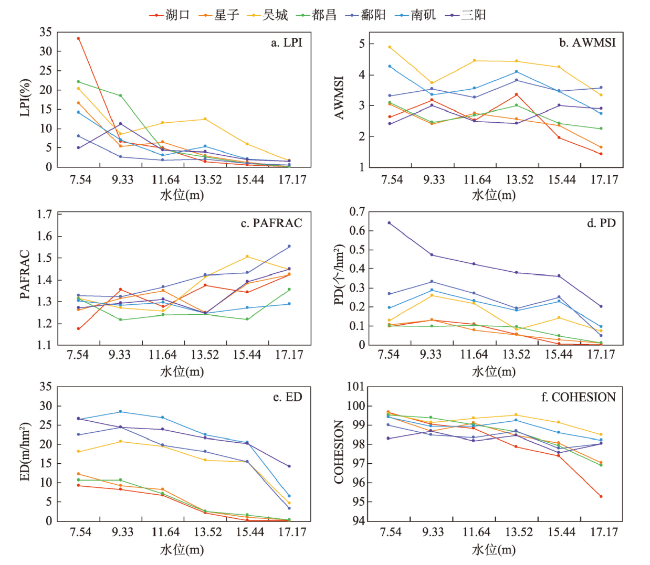
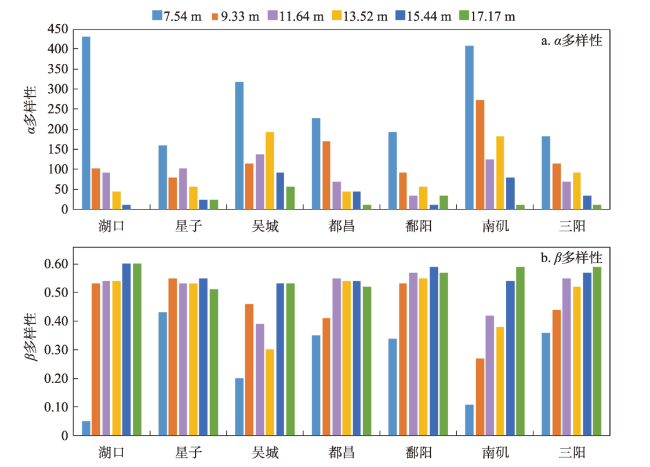
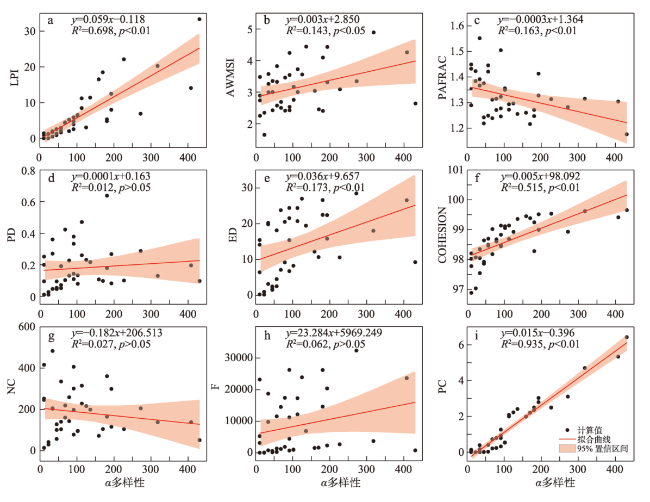
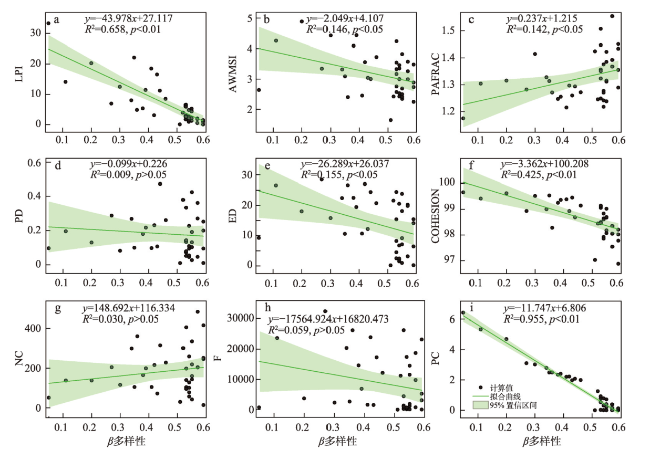
在全球景观破碎化不断加剧的背景下,剖析景观连通度对生物多样性的影响机制对恢复生态过程和保护生物多样性具有深远意义。本文以鄱阳湖草洲为研究对象,综合考虑水位变化与物种扩散生态过程,在识别不同水位梯度下草洲分布格局的基础上,通过景观格局指数和基于图论的景观连通度指数相结合的方法,分析不同水位草洲景观结构连通度和功能连通度的动态变化,揭示草洲景观连通度对植物多样性的影响及其尺度效应,比较结构连通度与功能连通度对植物多样性格局的相对解释力。研究结果表明:① 鄱阳湖草洲呈现“水落滩出、水进草退”的景观格局特征,随着水位的增加,草洲同时面临水体的淹没效应和分割效应,面积萎缩和破碎化并存;② 随着水位的增加,草洲斑块面积不断减小、形状趋于简单、密度降低、破碎化加剧,景观结构连通度下降,难以连接的独立组分增加、斑块连通的可能性降低,景观功能连通度下降;③ 草洲景观连通度越高,景观尺度上的植物物种丰富度越高,斑块尺度上的植物群落相似性越大,功能连通度比结构连通度更加能解释植物多样性格局。本文研究结果可从景观连通度的新视角为湖泊地区景观格局优化和植物多样性保护提供参考。
张成 , 陈文波 , 黄芳芳 . 鄱阳湖草洲景观连通性对植物多样性的影响[J]. 地理学报, 2024 , 79(1) : 259 -278 . DOI: 10.11821/dlxb202401016
With the acceleration of global landscape fragmentation, it is of great significance to reveal the impacts of landscape connectivity on biodiversity in order to restore ecological process and protect biodiversity. Taking the grassland of Poyang Lake as research object and from the integrated perspective of combined water level change and species dispersal, this study firstly analyzed the dynamics of grassland structural and functional connectivity respectively after identifying the distribution pattern of grassland at different water levels using landscape pattern indices and the graph-based connectivity indices. Then, the effects of grassland connectivity on plant diversity and the scale effects were discovered by means of the linear regression model. Moreover, the multiple stepwise linear regression model was applied to compare the relative explanatory power of structural and functional connectivity to plant diversity pattern. The results are shown as follows: (1) The grassland in Poyang Lake exhibits a landscape characteristic of "expanding when water falls while shrinking when water rises". It faces both inundation and segmentation effects of water with water level change, and its shrinkage always coexists with landscape fragmentation. (2) With the increase of water level, the area of grassland patch continues to shrink, the shape tends to be simple, the density reduces, and the fragmentation intensifies, resulting in a decrease of grassland structural connectivity. Meanwhile, the number of components increases, the probability of connectivity reduces, so that grassland functional connectivity decreases significantly. (3) The higher the connectivity of grassland, the higher the richness of plant species at the landscape scale, and the higher the similarity of plant communities at the patch scale. Functional connectivity is more effective in explaining plant diversity pattern than structural connectivity does. This study can provide some references for landscape pattern optimization and plant diversity conservation in lake areas from a new perspective of landscape connectivity.
Key words: Poyang Lake; grassland; landscape connectivity; plant diversity; scale effect
表1 遥感影像数据信息Tab. 1 Information of remote sensing image data |
| 序号 | 卫星/传感器 | 影像获取时间 | 星子站水位(m) |
|---|---|---|---|
| 1 | Landsat7/ETM+ | 2008-01-01 | 7.54 |
| 2 | Landsat7/ETM+ | 2019-11-15 | 9.33 |
| 3 | Landsat7/ETM+ | 2010-11-06 | 11.64 |
| 4 | Landsat5/TM | 2001-10-20 | 13.52 |
| 5 | Landsat7/ETM+ | 2010-10-05 | 15.44 |
| 6 | Landsat7/ETM+ | 2007-07-25 | 17.17 |
| 7 | Landsat7/ETM+ | 2010-08-02 | 19.97 |
注:第7幅影像用于确定鄱阳湖自然湿地范围。 |
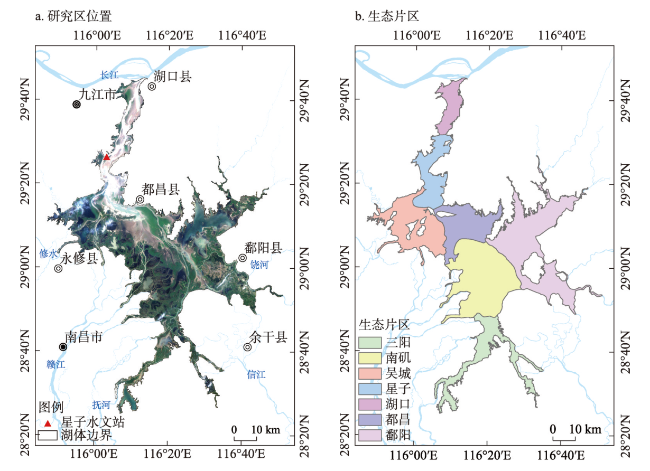
图2 研究区位置及生态片区Fig. 2 Spatial location of the study area (Poyang Lake) and its ecological zone |
表2 鄱阳湖各生态片区特征Tab. 2 Characteristics of each ecological zone of Poyang Lake |
| 区域名称 | 面积(km2) | 面积比(%) | 描述 |
|---|---|---|---|
| 湖口片区 | 165.01 | 5.16 | 屏峰至湖口河道型通江水体区 |
| 星子片区 | 289.58 | 9.06 | 松门山至屏峰河道型通江水体区 |
| 吴城片区 | 461.25 | 14.43 | 松门山以南湖泊型通江水体区 |
| 都昌片区 | 368.77 | 11.53 | 赣江北支与修河三角洲 |
| 鄱阳片区 | 810.60 | 25.35 | 赣江中支、南支河口三角洲 |
| 南矶片区 | 766.35 | 23.97 | 信江、饶河、通津河河口三角洲 |
| 三阳片区 | 335.83 | 10.50 | 抚河、信江河口河漫滩湿地区 |
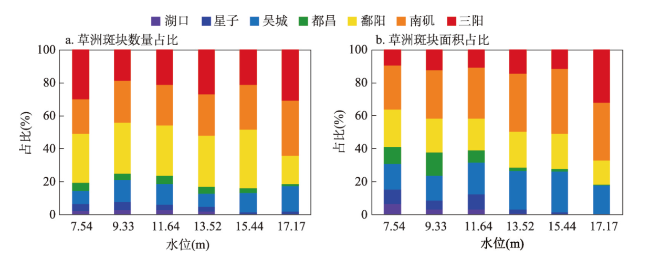
表3 鄱阳湖不同水位梯度下各生态片区草洲斑块的数量和面积Tab. 3 Number and area of grassland patches in each ecological zone of Poyang Lake at different water levels |
| 水位(m) | 草洲斑块 | 湖口 | 星子 | 吴城 | 都昌 | 鄱阳 | 南矶 | 三阳 | 合计 |
|---|---|---|---|---|---|---|---|---|---|
| 7.54 | 数量(个) | 16 | 30 | 58 | 38 | 214 | 149 | 217 | 722 |
| 面积(km2) | 119.33 | 157.79 | 277.50 | 192.11 | 407.19 | 488.17 | 171.52 | 1813.61 | |
| 9.33 | 数量(个) | 23 | 39 | 109 | 32 | 249 | 206 | 152 | 810 |
| 面积(km2) | 40.37 | 70.52 | 196.54 | 185.35 | 267.91 | 380.17 | 163.25 | 1304.11 | |
| 11.64 | 数量(个) | 18 | 24 | 86 | 34 | 210 | 169 | 144 | 685 |
| 面积(km2) | 34.80 | 96.04 | 208.73 | 81.42 | 205.71 | 332.21 | 114.37 | 1073.28 | |
| 13.52 | 数量(个) | 9 | 15 | 40 | 19 | 155 | 124 | 133 | 495 |
| 面积(km2) | 6.60 | 22.55 | 226.49 | 20.29 | 209.57 | 334.46 | 141.55 | 961.51 | |
| 15.44 | 数量(个) | 1 | 7 | 66 | 16 | 197 | 151 | 117 | 555 |
| 面积(km2) | 0.86 | 6.37 | 128.51 | 8.55 | 110.06 | 201.66 | 59.94 | 515.95 | |
| 17.17 | 数量(个) | 0 | 4 | 33 | 3 | 37 | 72 | 66 | 215 |
| 面积(km2) | 0.00 | 0.51 | 32.11 | 0.46 | 26.94 | 63.87 | 58.47 | 182.36 |
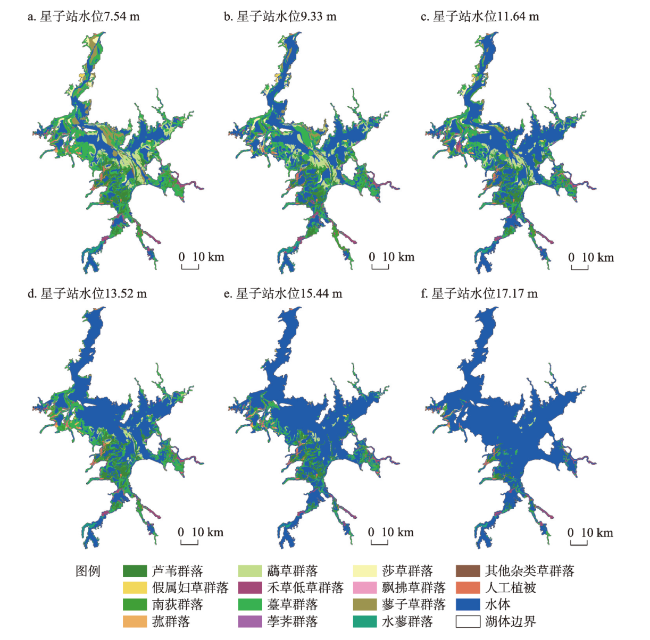
表4 鄱阳湖草洲植物群落分类Tab. 4 Classification of plant communities in Poyang Lake |
| 植被型 | 植被亚型 | 群系 |
|---|---|---|
| 草本沼泽植被型 | 禾草高草湿地亚型 | 芦苇群系、假鼠妇草群系、南荻群系、菰草群系、虉草群系 |
| 禾草低草湿地亚型 | 李氏禾群系、野古草群系、糠稷群系、白茅群系、稗草群系、狗尾草群系、狗牙根群系、假俭草群系、牛鞭草群系 | |
| 莎草湿地亚型 | 薹草群系、荸荠群系、莎草群系、飘拂草群系 | |
| 杂类草湿地亚型 | 香蒲群系、水烛群系、下江委陵菜群系、水田碎米荠群系、蓼子草群系、蚕茧蓼群系、酸模叶蓼群系、竹叶小蓼群系、丛枝蓼群系、疏花蓼群系、水蓼群系、齿果酸模群系、蒌蒿群系、细叶艾群系、芫荽菊群系、紫云英群系、半边莲群系 | |
| 人工植被 | 人工林 | 加拿大杨群系、乌桕群系、旱柳群系 |
表5 不同水位梯度下植物群落所占据的草洲斑块数量和面积Tab. 5 The number and area of grassland patch occupied by plant communities at different water levels |
| 植物群落类型 | 草洲斑块 | 水位(m) | |||||
|---|---|---|---|---|---|---|---|
| 7.54 | 9.33 | 11.64 | 13.52 | 15.44 | 17.17 | ||
| 芦苇群落 | 数量(个) | 199 | 296 | 291 | 219 | 250 | 87 |
| 面积(km2) | 197.82 | 148.83 | 137.31 | 166.47 | 104.54 | 34.35 | |
| 假鼠妇草群落 | 数量(个) | 3 | 3 | 3 | 5 | 3 | 0 |
| 面积(km2) | 7.26 | 5.89 | 4.25 | 1.86 | 1.74 | 0.00 | |
| 南荻群落 | 数量(个) | 114 | 174 | 165 | 184 | 164 | 52 |
| 面积(km2) | 98.57 | 82.80 | 78.19 | 89.23 | 55.21 | 15.00 | |
| 菰草群落 | 数量(个) | 27 | 36 | 36 | 23 | 36 | 7 |
| 面积(km2) | 10.19 | 7.11 | 8.46 | 10.32 | 7.62 | 2.81 | |
| 虉草群落 | 数量(个) | 133 | 206 | 189 | 157 | 95 | 8 |
| 面积(km2) | 210.06 | 176.46 | 109.55 | 53.20 | 20.75 | 4.32 | |
| 禾草低草群落 | 数量(个) | 116 | 134 | 119 | 114 | 121 | 59 |
| 面积(km2) | 64.03 | 63.47 | 54.43 | 51.72 | 36.36 | 28.49 | |
| 薹草群落 | 数量(个) | 695 | 1007 | 845 | 665 | 702 | 212 |
| 面积(km2) | 876.40 | 625.51 | 531.75 | 503.74 | 244.38 | 63.25 | |
| 荸荠群落 | 数量(个) | 3 | 3 | 3 | 3 | 2 | 0 |
| 面积(km2) | 3.99 | 2.68 | 2.12 | 1.52 | 0.60 | 0.00 | |
| 莎草群落 | 数量(个) | 20 | 36 | 31 | 23 | 8 | 2 |
| 面积(km2) | 59.44 | 41.90 | 32.74 | 5.47 | 2.43 | 1.19 | |
| 飘拂草群落 | 数量(个) | 3 | 3 | 3 | 3 | 3 | 0 |
| 面积(km2) | 1.11 | 0.88 | 1.02 | 0.94 | 0.59 | 0 | |
| 蓼子草群落 | 数量(个) | 86 | 127 | 95 | 39 | 12 | 3 |
| 面积(km2) | 204.16 | 84.23 | 53.98 | 6.33 | 2.79 | 0.08 | |
| 水蓼群落 | 数量(个) | 59 | 57 | 23 | 36 | 11 | 17 |
| 面积(km2) | 25.25 | 25.10 | 12.02 | 19.12 | 5.21 | 8.04 | |
| 其他杂类草群落 | 数量(个) | 37 | 36 | 38 | 40 | 11 | 7 |
| 面积(km2) | 9.89 | 5.95 | 5.89 | 5.22 | 1.51 | 1.22 | |
| 人工植被 | 数量(个) | 65 | 102 | 96 | 91 | 82 | 67 |
| 面积(km2) | 46.55 | 34.18 | 42.59 | 47.31 | 32.81 | 23.61 | |
| 合计 | 数量(个) | 1557 | 2217 | 1934 | 1599 | 1497 | 521 |
| 面积(km2) | 1813.61 | 1304.11 | 1073.28 | 961.51 | 515.95 | 182.36 | |
表6 植物多样性与草洲景观连通度指数的多元逐步线性回归分析Tab.6 Multiple stepwise linear regression analysis for plant diversity and grassland connectivity indices |
| 因变量 | 自变量 | 系数 | 标准化系数 | 标准差 | t值 | p值 | R2 |
|---|---|---|---|---|---|---|---|
| α多样性 | 常量 | -1404.297 | - | 742.285 | -1.892 | 0.037* | 0.951 |
| PC | 57.182 | 0.893 | 3.235 | 17.673 | <0.001** | ||
| NC | -0.091 | -0.101 | 0.033 | -2.741 | 0.009** | ||
| COHESION | 14.797 | 0.099 | 7.551 | 1.959 | 0.028* | ||
| β多样性 | 常量 | 0.541 | - | 0.008 | 67.987 | <0.001** | 0.975 |
| PC | -0.101 | -1.219 | 0.005 | -18.735 | 0.000** | ||
| LPI | 0.003 | 0.176 | 0.001 | 3.084 | 0.004** | ||
| ED | 0.003 | 0.188 | 0.001 | 5.176 | <0.001** |
注: *和**分别代表95%和99%的置信水平。 |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
[陈利顶, 刘雪华, 傅伯杰. 卧龙自然保护区大熊猫生境破碎化研究. 生态学报, 1999, 19(3): 291-297.]
|
| [10] |
|
| [11] |
[金奇豪, 汪倩旻, 李艺, 等. 中国沿海陆地自然保护区景观连接度评价. 地理学报, 2021, 76(11): 2830-2840.]
|
| [12] |
|
| [13] |
|
| [14] |
[刘世梁, 尹艺洁, 杨珏婕, 等. 漫湾库区景观破碎化对区域生境质量的影响. 生态学报, 2017, 37(2): 619-627.]
|
| [15] |
|
| [16] |
[陈春娣, 贾振毅, 吴胜军, 等. 基于文献计量法的中国景观连接度应用研究进展. 生态学报, 2017, 37(10): 3243-3255.]
|
| [17] |
[管超毅, 陈智, 黄乘明, 等. 广西黑叶猴栖息地景观格局破碎化分析及其对种群的影响. 生态学报, 2022, 42(3): 1203-1212.]
|
| [18] |
[李景霞, 付碧宏. 江苏盐城滨海湿地系统格局变化及其对丹顶鹤生境的影响. 遥感学报, 2021, 25(12): 2507-2519.]
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
[孙兴, 蒋小明, 宋高飞, 等. 青海湖浮游植物群落时空格局及其环境驱动因子. 湖泊科学, 2023, 35(2): 540-556.]
|
| [23] |
[张方方, 齐述华, 廖富强, 等. 鄱阳湖湿地出露草洲分布特征的遥感研究. 长江流域资源与环境, 2011, 20(11): 1361-1367.]
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
[陈利顶, 傅伯杰. 景观连接度的生态学意义及其应用. 生态学杂志, 1996, 15(4): 37-42.]
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
[刘瑞清, 李加林, 孙超, 等. 基于Sentinel-2遥感时间序列植被物候特征的盐城滨海湿地植被分类. 地理学报, 2021, 76(7): 1680-1692.]
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
[王伟, 周越, 田瑜, 等. 自然保护地生物多样性保护研究进展. 生物多样性, 2022, 30(10): 52-65.]
|
| [49] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}