
长期人类活动对浙江全新世植被的影响
|
王皓言(1999-), 男, 浙江海盐人, 硕士生, 研究方向为古生态学。E-mail: wanghaoyan000@qq.com |
收稿日期: 2022-11-01
修回日期: 2023-04-16
网络出版日期: 2023-05-27
基金资助
中国科学院战略性先导科技专项(XDB31030104)
中国科学院战略性先导科技专项(XDA2009000003)
浙江省自然科学基金项目(LY20D010002)
浙江省自然科学基金项目(LYD20D010003)
Long-term impacts of anthropogenic disturbances on Holocene vegetation in Zhejiang, China
Received date: 2022-11-01
Revised date: 2023-04-16
Online published: 2023-05-27
Supported by
Progress of Strategy Priority Research Program of Chinese Academy of Sciences(XDB31030104)
Progress of Strategy Priority Research Program of Chinese Academy of Sciences(XDA2009000003)
Zhejiang Natural Science Foundation(LY20D010002)
Zhejiang Natural Science Foundation(LYD20D010003)
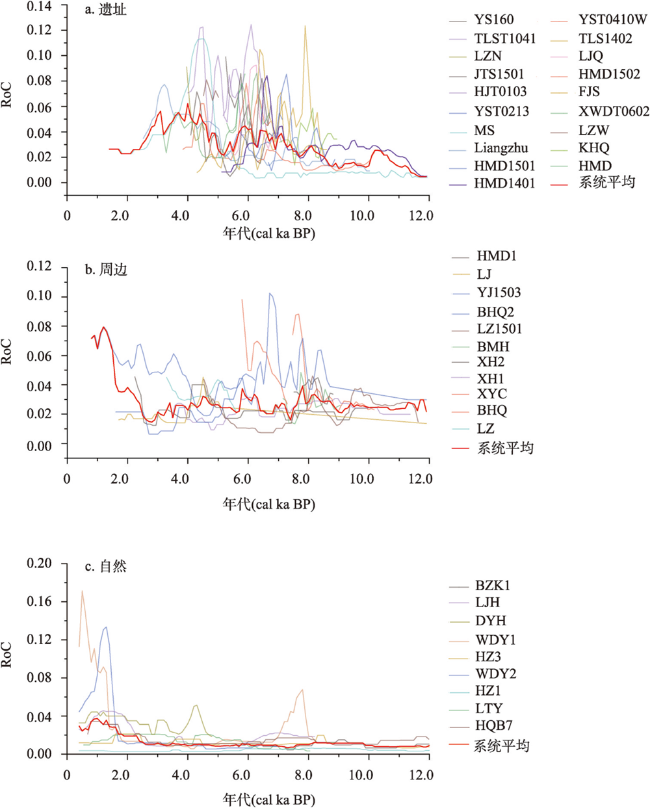
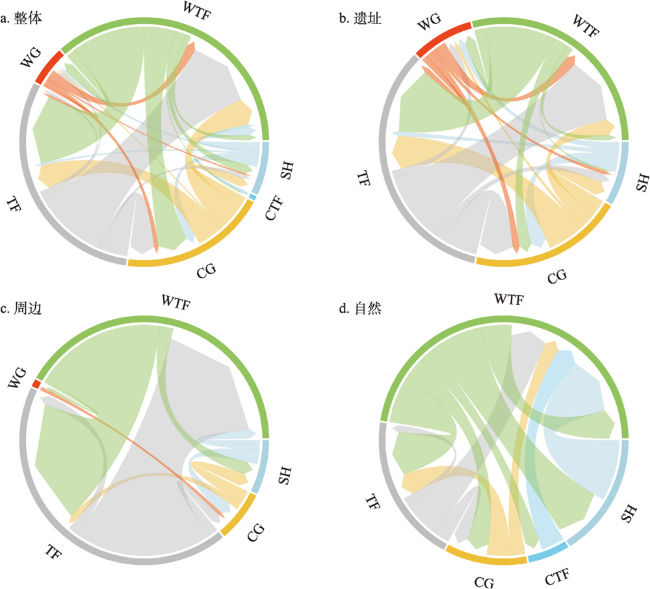
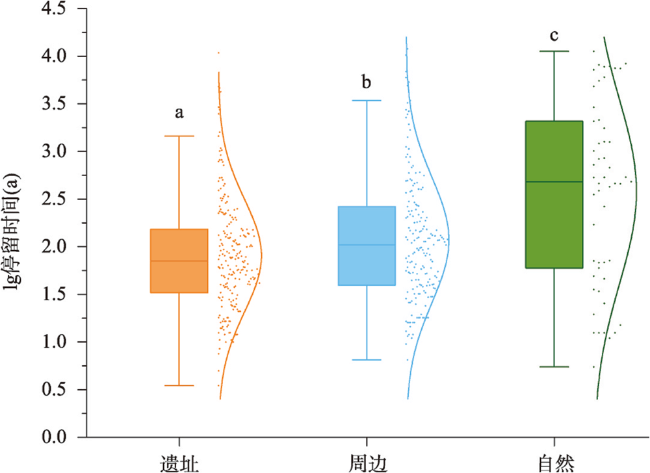
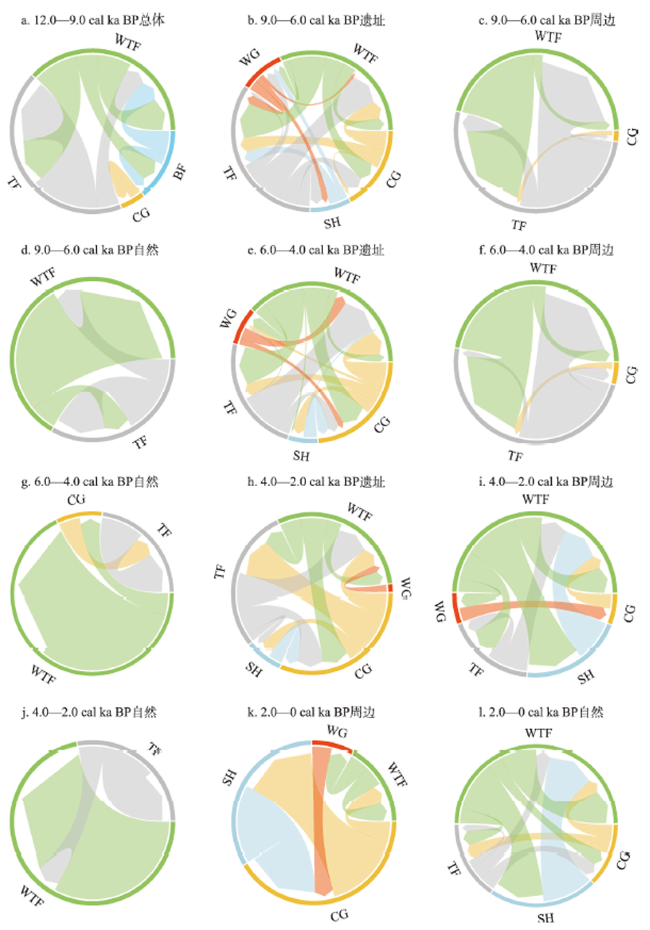
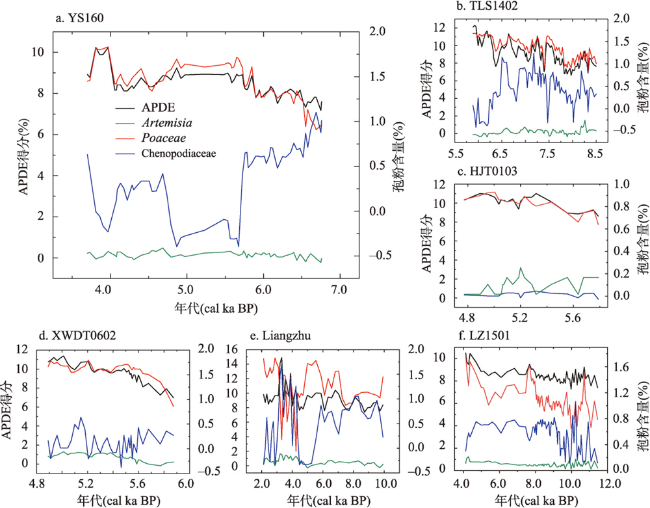
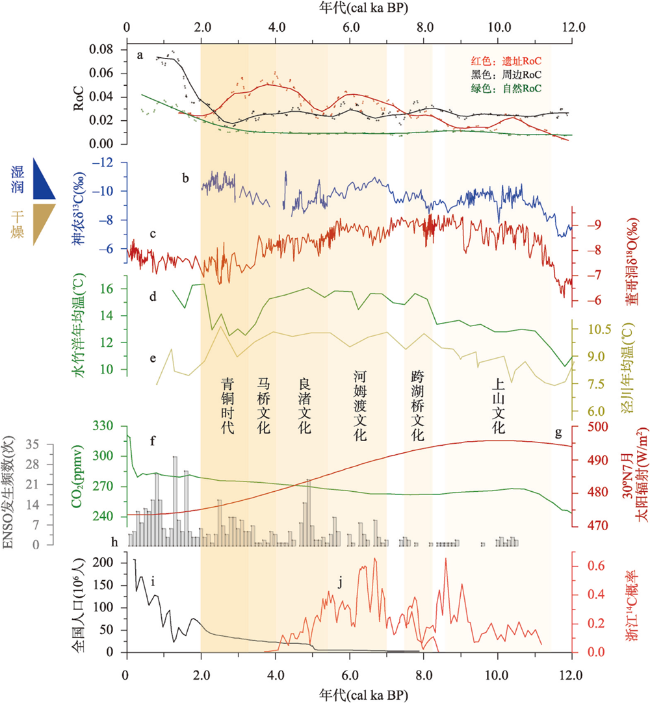
全新世人类活动逐渐增强,其对自然植被的干扰和改造也逐渐加深,在部分地区已然成为引起植被变化不可忽视的因素之一。考古结果证实,浙江地区在过去10 ka存在连续的文化序列,但全新世人类活动对区域植被的影响程度和范围尚不明确。本文收集、整理了浙江地区共40个样点的全新世孢粉记录,按照有无史前文化的发生将其分为遗址、遗址周边和自然3类数据,利用RRatepol程序包计算了各个样点的植被变化速率(RoC),同时使用孢粉生物群区化方法(Biomisation)重建了古植被变化过程。结果显示,在文化遗址,全新世RoC变化幅度较大且波动频繁,9.0—3.0 cal ka BP期间整体呈增加趋势;遗址周边的RoC变化幅度介于遗址和自然点位之间,在3.0 cal ka BP之后呈现迅速增加;自然点的RoC变幅最小且较为稳定,在2.0 cal ka BP后才出现明显升高。全新世期间人类活动引起的局部植被变化速率可能是自然植被变化的2~3倍,并且在3.0 cal ka BP后,人类活动对浙江古植被影响的空间范围开始明显扩张。人类活动导致的植被变化与自然植被变化存在一定的背离,新石器时代毁林开荒和种植农业的发展导致了浙江地区植被景观的破碎化。遗址点的生物群区停留时间明显低于遗址周边及自然点,说明全新世人类活动加速了区域植被成分变化的同时,也导致植被生态系统的弹性和恢复力显著降低。在浙江地区以孢粉进行的气候重建以及自然保护地体系建设实践中面临的植被保护和恢复问题,必须考虑长期的人类干扰历史以及植被变化历史格局的潜在影响。
王皓言 , 杨晹 , 周伯睿 , 李凯 , 廖梦娜 , 倪健 . 长期人类活动对浙江全新世植被的影响[J]. 地理学报, 2023 , 78(5) : 1153 -1175 . DOI: 10.11821/dlxb202305007
During the Holocene, human activities gradually intensified, resulting in significant interference and alteration of natural vegetation. Anthropogenic disturbance has been one of the most critical factors driving vegetation changes in some regions. The archaeological study illustrated that continuous Neolithic culture developed in Zhejiang during Holocene. However, the relationship between vegetation changes and human activities in this region remains unclear. Here, we collected 40 pollen records in Zhejiang and classified them into three groups, i.e., archaeological sites, peripheral sites and natural sites. After standardizing the chronology and taxonomy of those pollen data, we calculated the rate and pattern of vegetation changes (RoCs) by the R-Ratepol package and Biomisation. The results showed that the RoCs increased significantly at archaeological sites, with high variabilities during 9000-3000 cal a BP. The values of RoCs at peripheral sites were lower than that of archaeological sites but showed a dramatic increase after 3000 cal a BP. The RoCs at natural sites were the lowest and relatively stable before 2000 cal a BP; after 2000 cal a BP, it accelerated and outstripped that of archaeological and peripheral sites. The increased RoCs at both the peripheral and natural sites manifested that the spatial range of human impact on vegetation expanded significantly in the late Holocene. The vegetation change rates exerted by Neolithic human activities might be 2-3 times greater than that under the natural state. Furthermore, there was a certain deviation between the human-related and natural vegetation changes. The deforestation and cultivation of Neolithic ancestors led to the fragmentation of the terrestrial landscape. The residence time of biomes at archaeological sites was significantly lower than that in natural and peripheral sites. To sum up, long-term human activities in Zhejiang accelerated the regional vegetation changes and reduced the stability of terrestrial ecosystems. In pollen-based vegetation and climate reconstructions, and also in the practice of vegetation protection and restoration, the potential influence of long-term human activities must be considered.
Key words: vegetation change; human activities; Zhejiang; Holocene; pollen
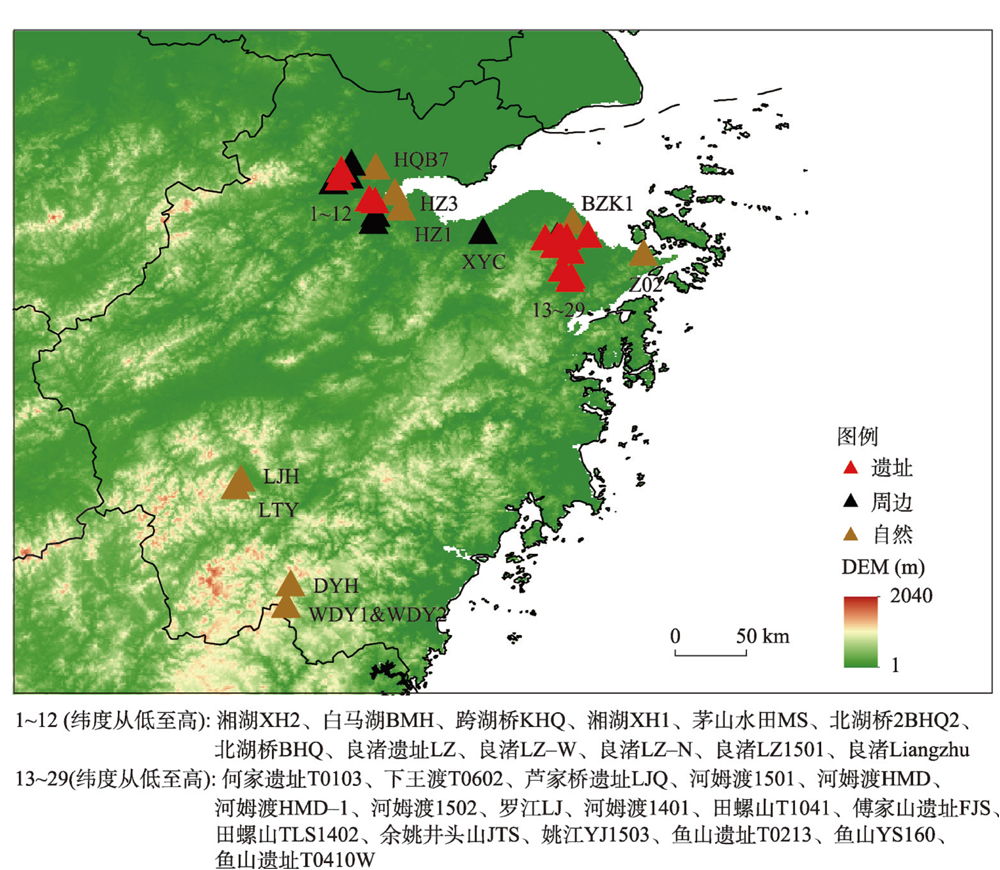
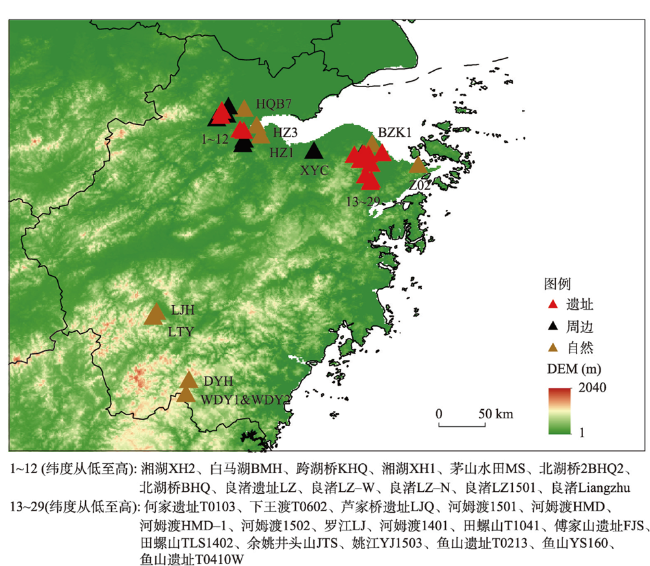
附表 本文采用的孢粉样点信息Appendix Table Sample sites information |
| 属性 | 地点 | 经度(°) | 纬度(°) | 类型 | 14C测年数量 | 时间跨度 (cal ka BP) | 文化阶段 | 海侵 | 数据 来源 |
|---|---|---|---|---|---|---|---|---|---|
| 遗址 | 跨湖桥KHQ | 120.2 | 30.3 | 遗址探方 | 11 | 6.6—8.9 | 跨湖桥 | 是 | [1] |
| 遗址 | 傅家山遗址FJS | 121.4 | 30.0 | 陆地钻孔 | 6 | 3.9—8.6 | 河姆渡 | 是 | [2] |
| 遗址 | 鱼山遗址YS160 | 121.6 | 30.0 | 陆地钻孔 | 4 | 3.7—6.7 | 河姆渡、良渚 | 是 | [2] |
| 遗址 | 鱼山遗址YST0213 | 121.6 | 30.0 | 遗址探方 | 8 | 4.4—7.1 | 河姆渡、良渚 | 是 | [3] |
| 遗址 | 鱼山遗址YST0410W | 121.6 | 30.0 | 遗址探方 | 7 | 3.4—6.6 | 河姆渡 | 是 | [2] |
| 遗址 | 河姆渡HMD | 121.3 | 30.0 | 河流相钻孔 | 6 | 2.5—6.7 | 河姆渡 | - | [4] |
| 遗址 | 河姆渡HMD1401 | 121.4 | 30.0 | 陆地钻孔 | 19 | 3.2—14.6 | 河姆渡 | 是 | [5] |
| 遗址 | 河姆渡HMD1501 | 121.4 | 29.9 | 陆地钻孔 | 5 | 6.2—8.7 | 河姆渡 | 是 | [6] |
| 遗址 | 河姆渡HMD1502 | 121.4 | 30.0 | 陆地钻孔 | 4 | 5.9—10.0 | 河姆渡 | 是 | [6] |
| 遗址 | 田螺山TLS1402 | 121.4 | 30.0 | 陆地钻孔 | 6 | 5.8—8.5 | 河姆渡 | 是 | [7] |
| 遗址 | 田螺山TLST1041 | 121.3 | 30.0 | 遗址探方 | 10 | 3.6—7.1 | 河姆渡、良渚 | - | [8] |
| 遗址 | 良渚遗址Liangzhu | 120.0 | 30.4 | 遗址探方 | 2 | 2.1—9.9 | 良渚 | - | [9] |
| 遗址 | 何家遗址HJT0103 | 121.4 | 30.0 | 遗址探方 | 4 | 4.7—5.7 | 河姆渡、良渚 | - | [10] |
| 遗址 | 芦家桥遗址LJQ | 121.4 | 29.8 | 陆地钻孔 | 5 | 4.9—6.7 | 河姆渡 | 是 | [2] |
| 遗址 | 余姚井头山JTS1501 | 121.4 | 30.0 | 陆地钻孔 | 5 | 4.8—6.5 | 跨湖桥、河姆渡 | 是 | [11] |
| 遗址 | 下王渡XWDT0602 | 121.4 | 29.8 | 遗址探方 | 7 | 4.8—5.8 | 河姆渡、良渚 | 是 | [12] |
| 遗址 | 茅山水田MS | 120.2 | 30.3 | 遗址探方 | 3+4OSL | 1.0—16.0 | 良渚 | - | [13] |
| 遗址 | 良渚遗址LZ-N | 120.0 | 30.4 | 遗址探方 | 7 | 3.5—4.5 | 良渚 | - | [14] |
| 遗址 | 良渚遗址LZ-W | 120.0 | 30.4 | 遗址探方 | 2 | 4.1—4.9 | 良渚 | - | [14] |
| 周边 | 姚江YJ1503 | 121.4 | 30.0 | 陆地钻孔 | 10 | 1.2—12.0 | - | 是 | [15] |
| 周边 | 良渚LZ1501 | 120.0 | 30.4 | 陆地钻孔 | 4 | 4.1—11.0 | - | 是 | [16] |
| 周边 | 白马湖BMH | 120.2 | 30.2 | 湖相钻孔 | 8 | 6.7—9.0 | - | 是 | [17] |
| 周边 | 湘湖XH2 | 120.2 | 30.1 | 湖相钻孔 | 7 | 7.1—8.7 | - | 是 | [17] |
| 周边 | 河姆渡HMD1 | 121.4 | 30.0 | 河流相钻孔 | 11 | 1.8—15.0 | - | - | [18] |
| 周边 | 新一村XYC | 120.9 | 30.1 | 陆地钻孔 | 4 | 5.3—7.3 | - | 是 | [19] |
| 周边 | 北湖桥BHQ | 120.0 | 30.4 | 河流相钻孔 | 4 | 7.0—10.0 | - | - | [20] |
| 周边 | 北湖桥2BHQ2 | 120.0 | 30.4 | 陆地钻孔 | 6 | 0.4—6.0 | - | 是 | [19] |
| 周边 | 良渚LZ | 120.1 | 30.5 | 陆地钻孔 | 5 | 2.8—5.5 | - | - | [21] |
| 周边 | 罗江LJ | 121.4 | 30.0 | 三角洲钻孔 | 4 | 1.3—13.0 | - | - | [22] |
| 周边 | 湘湖XH1 | 120.2 | 30.3 | 陆地钻孔 | 6 | 3.0—11.0 | - | - | [23] |
| 自然 | 杭州HQB7 | 120.2 | 30.5 | 河流相钻孔 | 4 | 0.2—29.0 | - | 是 | [24] |
| 自然 | 宁波Z02 | 121.9 | 29.9 | 河流相钻孔 | 5 | 0.7—13.0 | - | 是 | [25] |
| 自然 | 烂浆湖钻孔LJH | 119.3 | 28.5 | 泥炭剖面 | 2 | 0—8.0 | - | - | [26] |
| 自然 | 烂田砚泥炭LTY | 119.3 | 28.4 | 泥炭剖面 | 5 | 0.1—8.0 | - | - | [27] |
| 自然 | 望东洋WDY2 | 119.6 | 27.7 | 泥炭剖面 | 4 | 0—7.4 | - | - | [26] |
| 自然 | BZK1 | 121.5 | 30.1 | 河流相钻孔 | 3 | 4.7—30.0 | - | 是 | [28] |
| 自然 | 大仰湖DYH | 119.7 | 27.8 | 泥炭剖面 | 5 | 0—5.4 | - | - | [29] |
| 自然 | 三盈村HZ1 | 120.4 | 30.2 | 陆地钻孔 | 4OSL | 0—15.0 | - | - | [30] |
| 自然 | 下沙镇HZ3 | 120.3 | 30.3 | 陆地钻孔 | 4OSL | 0—13.0 | - | - | [30] |
| 自然 | 望东洋WDY1 | 119.6 | 27.7 | 泥炭剖面 | 7 | 0—8.3 | - | - | [31] |
特别感谢马春梅、李春海、叶玮等提供部分孢粉数据,及审稿专家和编辑部老师建设性的修改意见。
[1] Zong Y Q, Chen Z Y, Innes J B, et al. Fire and flood management of coastal swamp enabled first rice paddy cultivation in east China. Nature, 2007, 449(7161): 459-462.
[2] Tang Liang. Middle Holocene seawater intrusion and human's rapid response in the Yaojiang-Ningbo coastal plain, East China [D]. Shanghai: East China Normal University, 2019.
[唐亮. 姚江—宁波平原中全新世海水入侵及古人类活动的快速响应[D]. 上海: 华东师范大学, 2019.]
[3] He K Y, Lu H Y, Zheng Y F, et al. Middle-Holocene sea-level fluctuations interrupted the developing Hemudu culture in the lower Yangtze River, China. Quaternary Science Reviews, 2018, 188: 90-103.
[4] Liu Y, Sun Q L, Fan D D, et al. Pollen evidence to interpret the history of rice farming at the Hemudu site on the Ningshao coast, eastern China. Quaternary International, 2016, 426: 195-203.
[5] Huang Zhenhui. A study on the relationship between Holocene environment and human activities in the Yaojiang plain, Zhejiang province: A palynological analysis based on HMD1401 core [D]. Nanjing: Nanjing University, 2019.
[黄振辉. 浙江姚江平原全新世环境与人类活动关系研究[D]. 南京: 南京大学, 2019.]
[6] Ouyang X H, Hao X D, Zheng L B, et al. Early to mid-Holocene vegetation history, regional climate variability and human activity of the Ningshao Coastal Plain, eastern China: New evidence from pollen, freshwater algae and dinoflagellate cysts. Quaternary International, 2019, 528: 88-99.
[7] Ma Chunmei, Wang Bingling, Liu Zeyu, et al. human activity of pollen records during middle-late Holocene in Tianluoshan site, Ningshao Plain. Quaternary Research, 2018, 38(5): 1304-1312.
[马春梅, 王冰翎, 刘泽雨, 等. 浙江田螺山遗址区孢粉学记录的中全新世植被、环境和人类活动初步研究. 第四纪研究, 2018, 38(5): 1304-1312.]
[8] Li C H, Zheng Y F, Yu S Y, et al. Understanding the ecological background of rice agriculture on the Ningshao plain during the Neolithic Age: Pollen evidence from a buried paddy field at the Tianluoshan cultural site. Quaternary Science Reviews, 2012, 35: 131-138.
[9] Li Y Y, Wu J, Hou S F, et al. Palaeoecological records of environmental change and cultural development from the Liangzhu and Qujialing archaeological sites in the middle and lower reaches of the Yangtze River. Quaternary International, 2010, 227(1): 29-37.
[10] Li H Y, Sun J E, Ma C M, et al. Paleoenvironmental evolution and human activities at the Hejia Site on the Ningshao coastal plain in eastern China. Frontiers in Earth Science, 2021, 8: 609912. DOI:10.3389/feart.2020.609912.
[11] Liu Zeyu. Pollen records of vegetation, climate changes and human activities duing the mid-late Holocene from JTS1501 in Ningshao Plain [D]. Nanjing: Nanjing University, 2018.
[刘泽雨. 宁绍平原中晚全新世植被、环境与人类活动耦合关系[D]. 南京: 南京大学, 2018.]
[12] He K Y, Lu H Y, Li Y N, et al. Cultural response to Middle Holocene sea‐level fluctuations in eastern China: A multi‐proxy approach. Boreas, 2020, 49(1): 71-88.
[13] Jin Y X, Mo D W, Li Y Y, et al. Ecology and hydrology of early rice farming: geoarchaeological and palaeo-ecological evidence from the Late Holocene paddy field site at Maoshan, the Lower Yangtze. Archaeological and Anthropological Sciences, 2019, 11(5): 1851-1863.
[14] Wang X C, Mo D W, Li C H, et al. Environmental changes and human activities at a fortified site of the Liangzhu culture in eastern China: Evidence from pollen and charcoal records. Quaternary International, 2017, 438: 189-197.
[15] Liu Y, Deng L J, He J, et al. Early to middle Holocene rice cultivation in response to coastal environmental transitions along the South Hangzhou bay of eastern China. Palaeogeography, Palaeoclimatology, Palaeoecology, 2020, 555: 109872. DOI:10.1016/j.palaeo.2020.109872.
[16] Ling Guangjiu. The landscape evolution and its influence on Liangzhu culture in the Liangzhu since the Last Glacial Maximum [D]. Nanjing: Nanjing Normal University, 2021.
[凌光久. 末次冰盛期以来良渚地区地貌演化及其对良渚文化的影响[D]. 南京: 南京师范大学, 2021.]
[17] Pan Haochen. Vegetation changes and human activities during the Neolithic period in the Kuahuquai area of Zhejiang: Comparison of pollen records between natural cores and archaeological profiles [D]. Hefei: University of Science and Technology of China, 2020.
[潘浩晨. 浙江跨湖桥地区新石器时代植被变化及人类活动[D]. 合肥: 中国科学技术大学, 2020.]
[18] Li Chunhai, Tang Lingyu, Wan Hewen, et al. Vegetation and human activity in Yuyao (Zhejiang province) inferred from the sporo-pollen record since the Late Holocene. Acta Micropalaeontologica Sinica, 2009, 26(1): 48-56.
[李春海, 唐领余, 万和文, 等. 晚更新世以来浙江余姚地区植被变化及人类活动. 微体古生物学报, 2009, 26(1): 48-56.]
[19] Zhang Yunxia. Environmental and vegetation history variabilities archived by pollen-spore of the drill core Beihuqiao2 in Hangjiahu Plain during the mid-late Holocene, China [D]. Jinhua: Zhejiang Normal University, 2017.
[章云霞. 杭嘉湖平原高分辨率孢粉记录的中—晚全新世环境变化[D]. 金华: 浙江师范大学, 2017.]
[20] Lu Wenchen, Ye Wei. Characteristics of pollen assemblage and climate change in the Holocene at Borehole BHQ in Pingyao area, Zhejiang province. Journal of Palaeogeography, 2014, 16(5): 687-701.
[陆文晨, 叶玮. 浙江瓶窑BHQ孔全新统孢粉组合特征与气候变化. 古地理学报, 2014, 16(5): 687-701.]
[21] Liu Yan, Li Maotian, Sun Qianli, et al. Palaeoclimatic and environmental changes since the Mid-Holocene in the Hangzhou bay and their possible impacts on the evolution of Liangzhu Culture. Journal of Lake Sciences, 2014, 26(2): 322-330.
[刘演, 李茂田, 孙千里, 等. 中全新世以来杭州湾古气候、环境变迁及对良渚文化的可能影响. 湖泊科学, 2014, 26(2): 322-330.]
[22] Atahan P, Itzstein-Davey F, Taylor D, et al. Holocene-aged sedimentary records of environmental changes and early agriculture in the lower Yangtze, China. Quaternary Science Reviews, 2008, 27(5-6): 556-570.
[23] Gu Mingguang, Chen Zhongda, Lu Chengzhong, et al. Holocene sporopollen records in the Xianghu area, Zhejiang and their palaeoclimatic significance. China Geology, 2006(5): 1144-1148.
[顾明光, 陈忠大, 卢成忠, 等. 浙江湘湖地区全新世孢粉记录及其古气候意义. 中国地质, 2006(5): 1144-1148.]
[24] Gu Mingguang. Late Quaternary sediments and paleoenvironmental evolution on the northern bank of the Qiantang River. Geology in China, 2009, 36(2): 378-386.
[顾明光. 钱塘江北岸晚第四纪沉积与古环境演变. 中国地质, 2009, 36(2): 378-386.]
[25] Zhang Peixin. Late Quaternary sporopollen assemblages and sedimentary environment in the coastal plain of southeast Ningbo [D]. Chengdu: Chengdu University of Technology, 2018.
[张培新. 宁波东南沿海平原晚第四纪孢粉组合与沉积环境分析[D]. 成都: 成都理工大学, 2018.]
[26] Wen Zhenming, Ye Wei, Ma Chunmei, et al. Middle-late Holocene vegetation history and climate change implied by pollens from Wangdongyang, southern Zhejiang Province, east China. Acta Micropalaeontologica Sinica, 2018, 35(3): 260-272.
[温振明, 叶玮, 马春梅, 等. 浙江南部望东垟孢粉记录的中晚全新世植被演化历史与气候变化. 微体古生物学报, 2018, 35(3): 260-272.]
[27] Ma T, Tarasov P E, Zheng Z, et al. Pollen- and charcoal-based evidence for climatic and human impact on vegetation in the northern edge of Wuyi Mountains, China, during the last 8200 years. The Holocene, 2016, 26(10): 1616-1626.
[28] Lin Qinglong, Lin Nan, Ma Hongjie. Late Quaternary sediments and paleoenvironmental evolution in Cixi, Hangzhou bay south coast area. Science Technology and Engineering, 2017, 17(9): 1-8.
[林清龙, 林楠, 马宏杰. 杭州湾南岸慈溪地区晚第四纪沉积与古环境演化. 科学技术与工程, 2017, 17(9): 1-8.]
[29] Wang H Y, Liao M N, Li K, et al. Liangzhu culture recessed earlier than expected under a variable climate in the mid-late Holocene. Palaeogeography, Palaeoclimatology, Palaeoecology. Under Review.
[30] Liu Jingwei, Zhao Shujun, Cheng Jie, et al. A study of vegetation and climate evolution since the Holocene near the banks of the Qiantang River in Hangzhou Bay. Earth Science Frontiers, 2007, 14(5): 235-245.
[刘静伟, 赵淑君, 程捷, 等. 杭州湾钱塘江两岸全新世以来的古植被及古气候研究. 地学前缘, 2007, 14(5): 235-245.]
[31] Zhao L, Ma C M, Wen Z M, et al. Vegetation dynamics and their response to Holocene climate change derived from multi-proxy records from Wangdongyang peat bog in southeast China. Vegetation History and Archaeobotany, 2021, 31(3): 247-260.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
[ 李宜垠, 周力平, 崔海亭. 人类活动的孢粉指示体. 科学通报, 2008, 53(9): 991-1002.]
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
[ 吕厚远. 中国史前农业起源演化研究新方法与新进展. 中国科学: 地球科学, 2018, 48(2): 181-199.]
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
[ 吴立, 朱诚, 郑朝贵, 等. 全新世以来浙江地区史前文化对环境变化的响应. 地理学报, 2012, 67(7): 903-916.]
|
| [16] |
|
| [17] |
[ 陈海燕, 徐德宇, 廖梦娜, 等. 中国现代花粉数据集. 植物生态学报, 2021, 45(7): 799-808.]
|
| [18] |
|
| [19] |
|
| [20] |
[ 潘浩晨. 浙江跨湖桥地区新石器时代植被变化及人类活动[D]. 合肥: 中国科学技术大学, 2020.]
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
[ 李凯, 金伊丽, 谭斌, 等. 浙江景宁县大仰湖沼泽泥炭地形成过程及气候背景. 第四纪研究, 2019, 39(6): 1384-1392.]
|
| [28] |
|
| [29] |
|
| [30] |
[ 林清龙, 林楠, 马宏杰. 杭州湾南岸慈溪地区晚第四纪沉积与古环境演化. 科学技术与工程, 2017, 17(9): 1-8.]
|
| [31] |
[ 张培新. 宁波东南沿海平原晚第四纪孢粉组合与沉积环境分析[D]. 成都: 成都理工大学, 2018.]
|
| [32] |
|
| [33] |
[ 郑云飞, 蒋乐平. 上山遗址出土的古稻遗存及其意义. 考古, 2007(9): 19-25, 99, 2.]
|
| [34] |
|
| [35] |
[ 王凤, 曾蒙秀, 朱丽东, 等. 上山遗址区沉积过程与人类活动的关系. 沉积学报, 2021, 39(3): 723-738.]
|
| [36] |
|
| [37] |
[ 顾延生, 米雨川, 曾剑威, 等. 杭嘉湖平原腹地新石器早期夹砂陶片水稻植硅体化石发现及环境意义. 第四纪研究, 2021, 41(5): 1438-1444.]
|
| [38] |
[ 潘艳, 郑云飞, 陈淳. 跨湖桥遗址的人类生态位构建模式. 东南文化, 2013(6): 54-65.]
|
| [39] |
[ 顾明光, 陈忠大, 卢成忠, 等. 浙江湘湖地区全新世孢粉记录及其古气候意义. 中国地质, 2006, 33(5): 1144-1148.]
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
[ 白国柱. 长江下游地区夏商时期考古学文化研究[D]. 南京: 南京大学, 2017.]
|
| [45] |
[ 曹锦炎. 浙江出土商周青铜器初论. 东南文化, 1989(6): 104- 112, 231-235.]
|
| [46] |
[ 宋建. 马桥文化的分区和类型. 东南文化, 1999(6): 6-14.]
|
| [47] |
|
| [48] |
[ 马春梅, 王冰翎, 刘泽雨, 等. 浙江田螺山遗址区孢粉学记录的中全新世植被、环境和人类活动初步研究. 第四纪研究, 2018, 38(5): 1304-1312.]
|
| [49] |
|
| [50] |
|
| [51] |
[ 金凯, 王飞, 韩剑桥, 等. 1982—2015年中国气候变化和人类活动对植被NDVI变化的影响. 地理学报, 2020, 75(5): 961-974.]
|
| [52] |
[ 王娜, 许清海, 张生瑞, 等. 白洋淀地区晚冰期以来的气候和环境演变. 地理学报, 2022, 77(5): 1195-1210.]
|
| [53] |
|
| [54] |
|
| [55] |
[ 饶志国, 秦倩倩, 魏士凯, 等. 全新世温度研究回顾及对历史人地关系的启示. 地理学报, 2022, 77(5): 1169-1180.]
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
[ 张雅薇, 曾蒙秀, 章千惠, 等. 全新世以来钱塘江流域遗址时空分布特征及其与环境变化的关系. 长江流域资源与环境, 2022, 31(9): 2022-2034.]
|
| [65] |
[ 董广辉, 仇梦晗, 李若, 等. 探讨过去人地关系演变机制的“支点”概念模型. 地理学报, 2021, 76(1): 15-29.]
|
| [66] |
|
| [67] |
|
| [68] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}