
1963—2018年中国垂柳和榆树开花始期积温需求的时空变化
|
陶泽兴(1989-), 男, 湖北十堰人, 博士后, 主要从事植被地理和物候学研究。E-mail: taozx.12s@igsnrr.ac.cn |
收稿日期: 2019-07-26
要求修回日期: 2020-04-06
网络出版日期: 2020-09-25
基金资助
国家重点研发计划(2018YFA0606103)
国家自然科学基金项目(41901014)
国家自然科学基金项目(41871032)
中国科学院地理科学与资源研究所可桢杰出青年学者计划(2018RC101)
中国科学院青年创新促进会(2018070)
版权
Spatio-temporal variations in the thermal requirement of the first flowering dates of Salix babylonica and Ulmus pumila in China during 1963-2018
Received date: 2019-07-26
Request revised date: 2020-04-06
Online published: 2020-09-25
Supported by
National Key R&D Program of China(2018YFA0606103)
National Natural Science Foundation of China(41901014)
National Natural Science Foundation of China(41871032)
Outstanding Young Scholars from IGSNRR(2018RC101)
Youth Innovation Promotion Association of the CAS(2018070)
Copyright
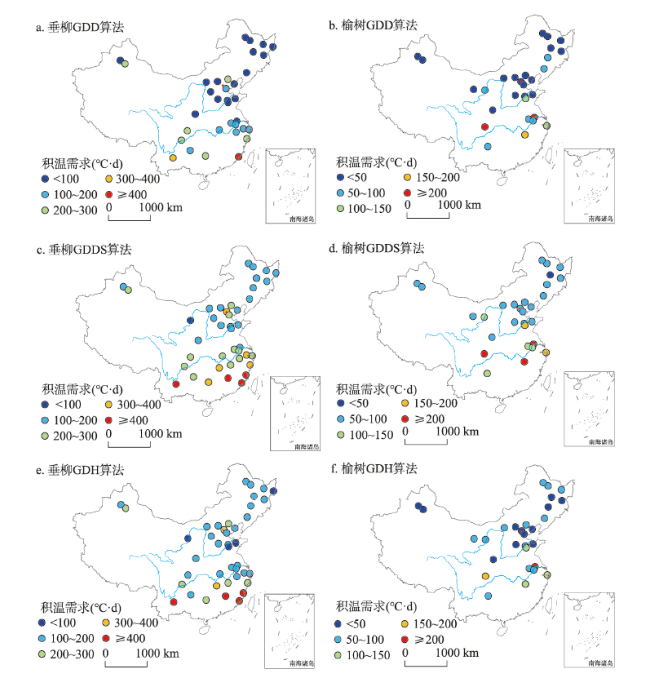
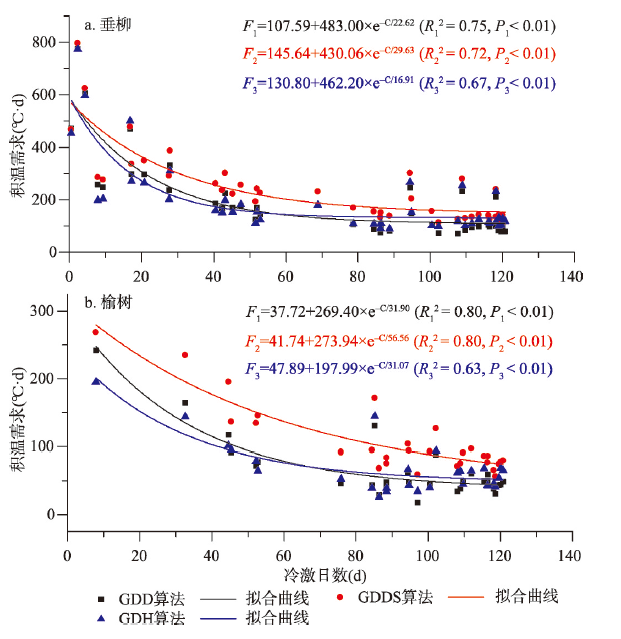
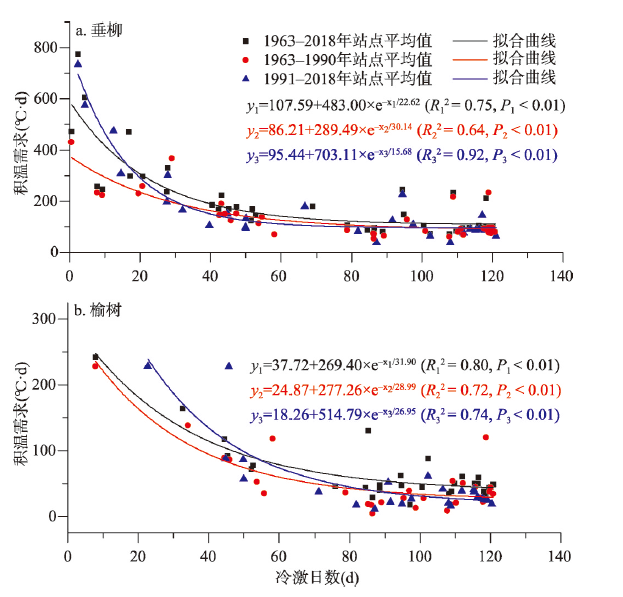
积温需求是决定北半球温带地区木本植物开花时间的主要因子。全球变暖使植物在冬季受到的冷激量减少,可能会改变植物开花的积温需求。气候变化导致的中国木本植物开花始期积温需求的时空变化仍不清楚。有鉴于此,本文基于“中国物候观测网”1963—2018年垂柳(Salix babylonica)和榆树(Ulmus pumila)开花始期数据,利用3种积温算法(GDD、GDDS和GDH)系统分析了两种植物开花积温需求的空间格局和在代表性站点的年际变化,构建了基于冷激日数模拟积温需求的模型。主要结论为:垂柳和榆树开花始期的积温需求在低纬度地区大于中纬度地区。站点多年平均积温需求与冷激日数呈显著的负指数关系,即随冷激日数增加,积温需求降低。时间上,3个典型站点(贵阳、西安和牡丹江)垂柳开花积温需求的变化趋势分别达到1.28~1.41 °C·d/a(P < 0.01)、1.63~1.89 °C·d/a(P < 0.01)和0.12~0.58 °C·d/a(仅GDD算法P < 0.05),榆树开花的积温需求在贵阳和西安同样显著增加,但在牡丹江变化不显著。冷激日数随时间减少是两个站点积温需求显著增加的主要原因。因牡丹江冬季气温低,冷激日数多且年际变化小,冷激对积温需求变化的影响不显著。基于时空耦合样本建立的冷激日数—积温需求模型对垂柳开花积温需求的模拟效果较好,R2达0.54~0.66。对榆树开花积温需求的模拟效果稍差(R2为0.33~0.64)。就不同算法而言,冷激日数对GDD算法得到的积温需求模拟效果更好。本文为量化植物开花积温需求的时空变化及在气候变化背景下的花期预测提供了重要的科学依据。
陶泽兴 , 葛全胜 , 王焕炯 . 1963—2018年中国垂柳和榆树开花始期积温需求的时空变化[J]. 地理学报, 2020 , 75(7) : 1451 -1464 . DOI: 10.11821/dlxb202007009
The forcing temperature in spring is the main factor that determines the flowering time of woody plants in the Northern Hemisphere. Global warming has reduced the number of chilling days in winter, which probably alters the thermal requirement of flowering. In the past 50 years, the spatio-temporal changes in the thermal requirement of spring phenology in China remain unclear. Based on the first flowering date (FFD) data of Salix babylonica and Ulmus pumila derived from China Phenological Observation Network during 1963-2018, we used three methods to calculate the thermal requirements of FFD and systematically analyzed their spatial and temporal patterns at representative sites. We also developed chilling days-thermal requirement models to quantitatively simulate the thermal requirement at each site in different years. The results showed that the thermal requirement of FFD exhibited a large spatial difference, with a relatively high value at low latitudes than that at middle latitudes. There was a significant negative exponential relationship between the average thermal requirement and chilling days across sites. The thermal requirements of FFD also changed over time. The trend in thermal requirement of Salix babylonica FFD in Guiyang, Xi'an and Mudanjiang reached 1.28-1.41 °C·d/a (P<0.01), 1.63-1.89 °C·d/a (P<0.01) and 0.12-0.58 °C·d/a (P<0.05) for the growing degree day method, respectively. The thermal requirement of Ulmus pumila FFD also increased significantly in Guiyang and Xi'an, but the trend in Mudanjiang was not significant. The decrease in the number of chilling days was the main reason for the increase in the thermal requirements. Due to the low winter temperature in Mudanjiang, the number of chilling days was large and had a small interannual variation, thus chilling days exerted no significant impact on the thermal requirement. The chilling days-thermal requirement model performed better in simulating the thermal requirement of Salix babylonica FFD, with R 2 of 0.54-0.66. In comparison with Salix babylonica, the model showed a relatively low precision in simulating the thermal requirement of Ulmus pumila FFD, with R 2 ranging from 0.33 to 0.64. Among the three methods, the thermal requirement could be better simulated by the growing degree days method compared with the growing degree days-sigmoid and growing degree hours methods. This study provides an important scientific basis for quantifying the spatio-temporal variation of the thermal requirement of flowering and for predicting the future flowering date of woody plants under the background of climate change.
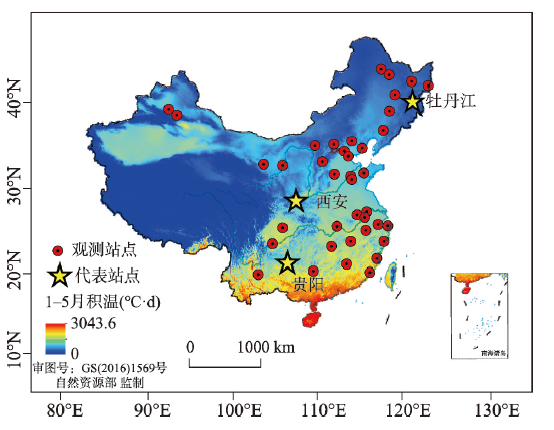
表1 物候观测站和对应气象观测站位置Tab. 1 The location of phenological observation sites and corresponding meteorological stations |
| 编号 | 物候 观测站 | 观测地点 | 垂柳观测起止年(年数) | 榆树观测起止年 (年数) | 气象站 (编号) | 纬度 (°N) | 经度 (°E) |
|---|---|---|---|---|---|---|---|
| 1 | 嫩江 | 嫩江农场 | 1975—1994(10) | 1974—1991(16) | 嫩江(50557) | 49.19 | 125.24 |
| 2 | 五大连池 | 龙镇农场 | 缺测 | 1974—1979(5) | 北安(50656) | 49.00 | 126.78 |
| 3 | 佳木斯 | 黑龙江农科院 佳木斯分院 | 1983—1988(5) | 1966—1996(22) | 佳木斯(50873) | 46.81 | 130.34 |
| 4 | 虎林 | 虎林市气象局 | 1983—1987(5) | 1964—1987(7) | 虎林(50983) | 45.77 | 132.97 |
| 5 | 哈尔滨 | 黑龙江省森林植物园 | 1963—1979(5) | 1963—2014(26) | 哈尔滨(50953) | 45.75 | 126.63 |
| 6 | 牡丹江 | 牡丹江农气试验站 | 1978—2018(42) | 1965—2018(42) | 牡丹江(54094) | 44.57 | 129.58 |
| 7 | 石河子 | 石河子大学 | 1984—1996(12) | 1963—1996(16) | 石河子(51356) | 44.35 | 85.95 |
| 8 | 长春 | 吉林省自然博物馆 | 2003—2018(16) | 1986—2018(25) | 长春(54161) | 43.88 | 125.35 |
| 9 | 乌鲁木齐 | 新疆林科院 | 1985—2018(5) | 1963—1990(8) | 乌鲁木齐(51463) | 43.75 | 87.60 |
| 10 | 沈阳 | 沈阳农业大学 | 1964—2018(23) | 1964—2018(26) | 沈阳(54342) | 41.80 | 123.38 |
| 11 | 承德 | 河北旅游职业学院 | 缺测 | 1974—1996(10) | 承德(54423) | 40.85 | 118.06 |
| 12 | 呼和浩特 | 内蒙古大学 | 1979—2012(12) | 1964—2004(12) | 呼和浩特(53463) | 40.80 | 111.68 |
| 13 | 张家口 | 张家口气象局 | 1974—1993(10) | 1974—1993(10) | 张家口(54401) | 40.78 | 114.90 |
| 14 | 北京 | 颐和园 | 1974—2018(6) | 1963—2012(43) | 北京(54511) | 40.02 | 116.33 |
| 15 | 秦皇岛 | 秦皇岛市地理学会 | 1980—1993(12) | 1980—1993(12) | 秦皇岛(54449) | 39.88 | 119.25 |
| 16 | 天津 | 园林绿化所 | 1980—1992(9) | 1980—1992(12) | 天津(54527) | 39.39 | 117.07 |
| 17 | 原平 | 原平县水利局 | 1977—1982(5) | 1976—1982(7) | 原平(53673) | 38.73 | 112.71 |
| 18 | 民勤 | 民勤沙生植物园 | 缺测 | 1974—1996(20) | 民勤(52681) | 38.63 | 103.08 |
| 19 | 银川 | 宁夏气象科研所 | 2003—2018(20) | 2006—2018(13) | 银川(53614) | 38.48 | 106.22 |
| 20 | 邢台 | 达活泉公园 | 1982—1996(15) | 1982—1996(15) | 邢台(53798) | 37.09 | 114.48 |
| 21 | 潍坊 | 潍坊市气象局 | 1967—1996(9) | 1985—1996(8) | 潍坊(54843) | 36.69 | 119.08 |
| 22 | 济南 | 山东省科学院 | 1965—2018(5) | 1963—2018(8) | 济南(54823) | 36.65 | 117.04 |
| 23 | 泰安 | 山东农业大学 | 1963—1985(12) | 1963—1981(6) | 泰安(54827) | 36.17 | 117.10 |
| 24 | 西安 | 西安植物园 | 1964—2018(39) | 1964—2015(31) | 泾河(57131) | 34.22 | 108.97 |
| 25 | 南京 | 九华山公园 | 1987—2017(17) | 1987—2018(10) | 南京(58238) | 32.04 | 118.78 |
| 26 | 合肥 | 合肥师范学院 | 1965—2018(37) | 1965—2018(35) | 合肥(58321) | 31.83 | 117.25 |
| 27 | 芜湖 | 安徽师范大学 | 1963—1996(18) | 1963—1996(14) | 芜湖(58334) | 31.28 | 118.38 |
| 28 | 武汉 | 狮子山 | 1963—1981(6) | 缺测 | 武汉(57494) | 30.52 | 114.31 |
| 29 | 杭州 | 杭州植物园 | 1963—1983(9) | 缺测 | 杭州(58457) | 30.25 | 120.12 |
| 30 | 宁波 | 宁波农业科学研究院 | 1968—1996(25) | 1981—1989(6) | 鄞县(58562) | 29.85 | 121.62 |
| 31 | 屯溪 | 黄山学院 | 1982—1996(14) | 缺测 | 屯溪(58531) | 29.69 | 118.29 |
| 32 | 南昌 | 江西农业大学 | 2008—2018(9) | 1985—1991(7) | 南昌(58606) | 28.77 | 115.83 |
| 33 | 长沙 | 中南林业科技大学 | 2007—2019(10) | 缺测 | 长沙(57679) | 28.20 | 113.07 |
| 34 | 温州 | 温州科技职业学院 | 1966—1974(9) | 缺测 | 温州(58659) | 27.98 | 120.63 |
| 35 | 贵阳 | 贵州大学 | 1963—2018(30) | 1963—2018(32) | 贵阳(57816) | 26.42 | 106.67 |
| 36 | 福州 | 福州农气试验站 | 2003—2018(10) | 缺测 | 福州(58847) | 26.08 | 119.33 |
| 37 | 桂林 | 桂林植物园 | 1964—2015(25) | 缺测 | 桂林(57957) | 25.18 | 110.20 |
| 38 | 昆明 | 昆明植物园 | 1963—2017(19) | 缺测 | 昆明(56778) | 25.04 | 102.73 |
| 39 | 厦门 | 厦门大学 | 1964—1988(8) | 缺测 | 厦门(59134) | 24.44 | 118.10 |
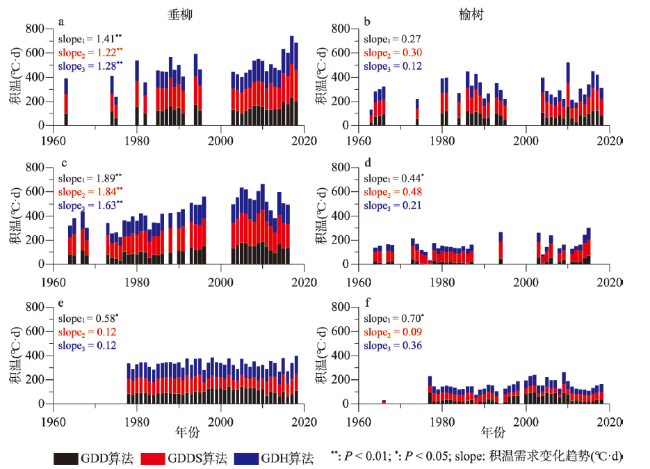
图4 1963—2018年垂柳和榆树开花始期积温需求在贵阳(a、b)、西安(c、d)和牡丹江(e、f)的年际变化注:slope1、slope2和slope3分别为GDD、GDDS和GDH算法的积温需求变化趋势。 Fig. 4 Interannual variation of thermal requirements for the first flowering dates of Salix babylonica and Ulmus pumila at Guiyang (a, b), Xi'an (c, d) and Mudanjiang (e, f) from 1963 to 2018 |
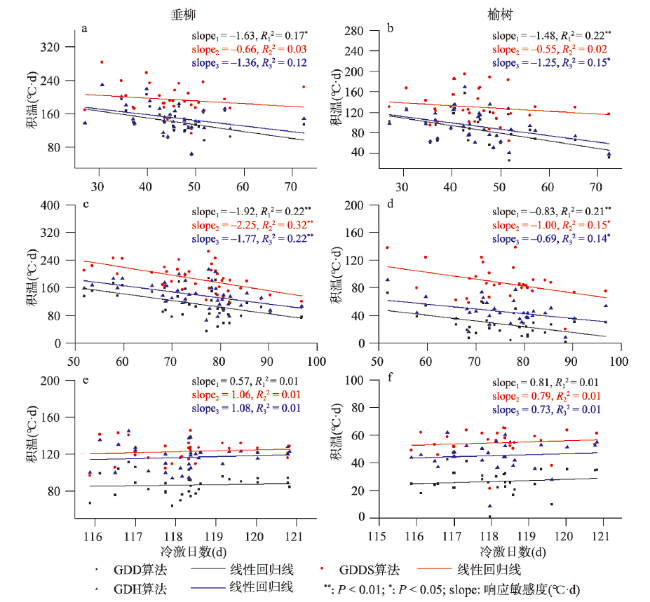
图5 贵阳(a、b)、西安(c、d)和牡丹江(e、f)垂柳和榆树开花始期的积温需求与冷激日数的关系注:slope1、slope2和slope3分别为GDD、GDDS和GDH算法的积温需求变化趋势。 Fig. 5 Relationship between the thermal requirements and chilling days for the first flowering dates of Salix babylonica and Ulmus pumila in Guiyang (a, b), Xi'an (c, d) and Mudanjiang (e, f) |
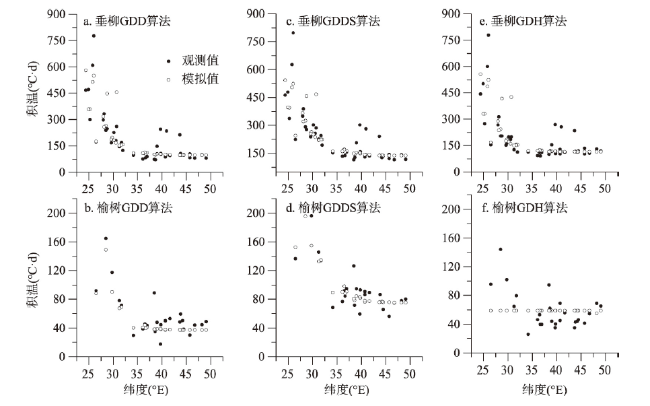
表2 基于时空耦合样本的冷激日数—积温需求模型公式Tab. 2 The equations of chilling day-thermal requirement models based on spatio-temporal mixed sample |
| 物种 | 积温需求算法 | 公式 | R2 | RMSE(°C·d) | P |
|---|---|---|---|---|---|
| 垂柳 | GDD | F=92.17+502.79×e-C/24.29 | 0.66 | 84.09 | < 0.01 |
| GDDS | F=122.47+428.79×e-C/34.99 | 0.60 | 85.09 | < 0.01 | |
| GDH | F=110.98+461.55×e-C/20.43 | 0.54 | 94.00 | < 0.01 | |
| 榆树 | GDD | F=36.30+774.01×e-C/15.89 | 0.64 | 29.75 | < 0.01 |
| GDDS | F=68.27+363.21×e-C/30.04 | 0.40 | 33.01 | < 0.01 | |
| GDH | F=29.82+132.32×e-C/47.74 | 0.33 | 32.32 | < 0.01 |
| [1] |
|
| [2] |
|
| [3] |
[ 陶泽兴, 仲舒颖, 葛全胜, 等. 1963—2012年中国主要木本植物花期长度时空变化. 地理学报, 2017,72(1):53-63.]
|
| [4] |
[ 张明庆, 杨国栋, 范振涛, 等. 北京地区主要致敏花粉树木花期的预报. 环境与健康杂志, 2008,25(3):262-263.]
|
| [5] |
|
| [6] |
[ 董海涛, 谭丽静, 刘洪林, 等. 基于积温模型丹东地区桃树盛花期预测研究. 气象与环境学报, 2018,34(1):99-105.]
|
| [7] |
IPCC. Summary for policymakers//Global Warming of 1.5°C. An IPCC Special Report on the impacts of global warming of 1.5 °C above pre-industrial levels and related global greenhouse gas emission pathways, in the context of strengthening the global response to the threat of climate change, sustainable development, and efforts to eradicate poverty. World Meteorological Organization, Geneva, Switzerland, 2018: 1-32.
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
[ 徐韵佳, 仲舒颖, 戴君虎, 等. 1978—2014年牡丹江地区植物花期变化及模型模拟. 地理研究, 2017,36(4):779-789.]
|
| [12] |
[ 张爱英, 王焕炯, 戴君虎, 等. 物候模型在北京观赏植物开花期预测中的适用性. 应用气象学报, 2014,25(4):483-492.]
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
[ 宛敏渭, 刘秀珍. 中国物候观测方法. 北京: 科学出版社, 1979: 1-24.]
|
| [27] |
[ 郑景云, 卞娟娟. 类型变更的相邻气象观测站的日气温资料整合. 地理研究, 2012,31(4):579-588.]
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
[ 戴君虎, 陶泽兴, 王焕炯, 等. 物候模型在气候定量重建中的应用. 第四纪研究, 2016,36(3):702-710.]
|
| [39] |
|
| [40] |
|
| [41] |
[ 仲舒颖, 葛全胜, 戴君虎, 等. 中国典型观赏植物花期模型建立及过去花期变化模拟. 资源科学, 2017,39(11):2116-2129.]
|
| [42] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}