
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
黄土高原山地森林群落植物区系特征与地理格局
[赵鸣飞1, 2  , 王宇航
, 王宇航1, 2 , 邢开雄1, 2 , 康慕谊1, 2 , 刘全儒3 , 李秋颐2 , 黄永梅2 ]
, 王宇航, 刘全儒|
|


作者简介:赵鸣飞, 博士研究生, 研究方向为植被生态学。E-mail: landscaper.mf@gmail.com
本文以样地数据为基础,对黄土高原内11座山地森林群落的区系组成、联系、格局以及与气候因子的关系等方面进行了较为系统的分析,结果表明:(1) 研究区内森林群落物种丰富,353个样地共调查到维管植物108科473属1222种,其中被子植物93科447属1179种,裸子植物4科7属9种,蕨类植物11科19属34种。(2) 按区域和局地两个尺度统计显示,研究区内科、属分布区类型以“温带分布”特别是“北温带分布”为主,此特征在群落尺度上尤显突出。(3) 非度量多维度标度排序 (NMDS) 第一轴主要揭示各山地森林分布的典型海拔变化范围,第二轴则展示出各山地的地理纬度位置关系;系统聚类树同样体现出各山地间基于区系组成特征的地理空间联系与南北位置关系,并将各山地依区系组成特征划分为4组;经比较山地间最大Sørenson相似性系数知,贺兰山因地处区域西北边缘而与其他山地间区系联系最弱,反之太岳山与其他山地区系联系最强。(4) R/T值 (热带属与温带属比值) 线性回归模型解释了42.1%的数据变异,并揭示该值随年均温升高呈显著上升趋势,随着海拔升高则有显著下降趋势,但与年均降水量之间并无显著趋势。
, WANG Yuhang, LIU QuanruBased on the sampling data, we analyzed and revealed the floristic characteristics and the relationships of floristic composition between and among 11 mountains on the Loess Plateau. The results indicate that: (1) The floristic composition in forest communities in this region is abundant. Among the 353 plots, 1222 species of vascular plants, belonging to 473 genera of 108 families, were discovered. In those species, 1179 species of 447 genera of 93 families are Angyospermae, 9 species of 7 genera of 4 families are Gymnospermae, and 34 species of 19 genera of 11 families are Pteridophyta. (2) On the regional and local scales, the dominant distribution types of floristic elements at the family and genus levels are temperate origin, especially the north temperate origin. This characteristic is unique on the scale of community level. (3) The first axis of NMDS ordination revealed typical variation range of elevational gradient. The second axis displayed the latitudinal relationship between and among different mountains. The whole study area can be divided into 4 groups with different levels of floristic linkage by hierarchical clustering. Through the comparison of maximum Sørenson similarity coefficient, Helan Mountain, located at the northwest edge of the region, had the weakest relation with other mountains, whereas Taiyue Mountain had the strongest relation with all of the other mountains. (4) Multiple linear regression analysis accounted for 42.1% of the variation within the R/T value (ratio of the tropical to the temperate elements), revealing a significant trend that the R/T value in the region increased at sites with high annual average temperature, whereas it decreased at sites with high elevation. However, no significant tendency of the R/T value with annual precipitation was found in the same analysis.
植物区系是植物类群在一定区域自然地理和历史条件综合作用下发展演化的结果, 作为群落结构的基础, 不仅能够间接、稳定地反映其所处地区的气候、土壤等自然条件特点[1, 2], 而且也是揭示区域植物群落内在性质和规律的重要特征指标。生态过渡带 (Ecological transitional area) 多指大尺度气候生物地理带之间的过渡区域, 虽不能完全等同于生态交错带 (Ecotone), 但往往具有生态交错带的许多特性, 如物种多样性较周边丰富、对环境变化具有敏感性和生态脆弱性等[3]。在生态过渡带, 物种与环境之间、物种与物种之间以及物种内部各个种群之间的相互作用往往有着鲜明的生态地理规律性。因此, 探讨过渡带区域植物群落的区系组成及其内部联系和分布特点等, 无疑具有重要的科学理论意义和现实资源保护和利用价值。
黄土高原地处我国温带季风气候区与温带大陆性气候区的交汇处, 气候具明显的半湿润向半干旱过渡特征, 是一个相对独立的空间自然地理单元和典型的生态过渡带。该区域的植物区系在演化历史上, 特别是气候频繁剧烈变化的最近历史时期 (如自第四纪更新世) 以来, 一直是欧亚温带植物区系与中国南方热带— 亚热带植物区系之间物种迁移扩散的重要场所[4]。我国学者曾在贺兰山[5]、太岳山[6]、关帝山[7]、芦芽山[8]、六盘山[9]、大青山[10]、子午岭[11, 12]、中条山[13, 14]等典型山地相继开展了有关植物区系的实地调查研究工作, 为黄土高原植物区系研究积累了重要的本底资料。但以往研究多以文献记录结合线路标本调查等传统方法为主要手段, 研究区多仅局限于黄土高原的一部分, 统计定量分析亦不足, 导致研究结果仍停留在以描述性比较为主的原初阶段。本研究采用系统取样法, 以在全区域内山地地区开展的较为全面的野外样方调查数据为基础, 尝试在局地和区域两种尺度上揭示黄土高原山地主要森林植物群落的区系组成和性质, 以及研究区内不同区域间的以此森林植物区系为基础的纽带联系与分异规律, 为黄土高原山地森林植被的切实保育与可持续利用等提供科学指导和决策参考依据。
黄土高原大多指位于北纬34° ~40° , 东经103° ~114° 之间, 包括太行山以西、日月山以东、秦岭以北、阴山以南, 面积约63´ 104 km2的广阔地区[15]。黄土高原属一个相对独立的地貌单元, 平均海拔800~1200 m, 其内部分基岩裸露的山地海拔可达1300~1700 m, 不乏一些海拔逾2000 m的山峰。除少数石质山地外, 黄土层堆积平均厚达50~80 m。气候以季风性向大陆性过渡为主, 年均温6~14 oC, 年均降水量300~800 mm, 从东南向西北递减, 对应的气候带依次为湿润半湿润暖温带、半湿润半干旱温带、干旱半干旱温带[15, 16, 17]。
本研究的采样区域集中于人为干扰较少、森林植被保存较为完好的11座山地 (图4)。这些山地覆盖了黄土高原绝大部分地理区域与生境梯度, 其年均温梯度范围达15 oC (变动在-1~14 ° C间), 年均降水量梯度范围达500 mm (变动在300~800 mm间)。土壤类型主要为褐土类 (包括淋溶褐土、灰褐土和石灰性褐土等各亚类) 和山地棕壤; 代表性林型主要包括寒温性的常绿或落叶针叶林, 如青海云杉 (Picea crassifolia) 林、华北落叶松 (Larix principis-rupprechtii) 林、青杄 (Picea wilsonii) 林; 温性常绿针叶林, 如油松 (Pinus tabulaeformis) 林; (温性) 落叶阔叶林, 如辽东栎 (Quercus wutaishanica) 林、白桦 (Betula platyphylla) 林、红桦 (Betula albo-sinensis) 林、山杨 (Populus davidiana) 林、杂木林, 以及两类森林过渡区形成的各种针阔混交林等。
参考群落调查相关技术规范[18], 2011-2013年间, 于植物生长旺季 (每年6-8月) 对各山地典型地段的代表性群落进行系统取样调查。样地布设兼顾局地生境与地形, 并依海拔梯度设置。通常从出现天然或半天然森林分布的低海拔处开始, 高程每升高100 m设置20 m× 30 m样地 (划分为6个10 m× 10 m样格) 2~3个。在此600 m2样方内进行乔木树种的每木调查 (胸径大于3 cm者), 记录其种名、胸径、树高和冠幅; 同时记录出现在样地内灌木层和草本层的全部物种, 并依《中国植物志》确定分类群所属单元。再选择处于对角位置的两个10 m× 10 m样格调查其内灌木的基径、高度、盖度和丛数等, 并在每样格中间位置取1 m× 1 m样方统计其内草本植物的盖度、高度和株数等特征。样地所处的地理位置信息以及其坡度、坡向等地形因子亦同时作详细记录。不同山地的取样强度, 依据其森林分布的海拔范围、面积、群落类型复杂程度等因素决定。由此共获得群落调查样方205个, 另外包括2005年65个10 m× 10 m贺兰山样地数据和2003-2004年间83个20 m× 20 m中条山样地数据 (表1), 取样方法与上面介绍基本一致。
| 表1 研究区山地及样地简况 Tab. 1 General features of 353 forest plots sampled from 11 mountains on the Loess Plateau |
年均温与年降水量数据获取自WorldClim数据库 (1.4版, http://www.progressingeography.com/richhtml/0375-5444/html_resources/www.world-clim.org)。
2.3.1 非度量多维标度排序 排序采用非度量多维标度法 (Nonmetric Multi-Dimensional Scaling, NMDS)。为消除部分山地由于取样面积不同所带来的偏差, 本研究采用了不同山地植物属的“ 区系谱” 数据 (Floristic Spectrum)[19]作为NMDS的原始矩阵。
2.3.2 聚类分析 以各山地植物属的出现频率作为原始数据矩阵进行系统聚类, 两山地之间的区系关系强度以Bray-Curtis距离度量[20]。组间聚合策略采用Ward离差平方和法, 并通过多响应置换过程分析 (Multi-response Permutation Procedures, MRPP) 检验分组结果的合理性是否显著[21]。
2.3.3 相似性系数计算 以各山地植物属出现与否的二元矩阵为基础, 采用Sø renson相似性系数度量山地之间的区系相似性程度, 公式如下:
式中, Sc为Sø renson系数, c为两地共有种数, a为甲地独有种个数, b为乙地独有种个数, 计算时依照区系分析惯例剔除掉世界分布属。该区系相似性指数因符合统计学规律而被广泛使用[22]。
2.3.4 R/T值计算与回归分析 计算每个样地热带区系成分 (Trop.) 与温带区系成分 (Temp.) 之比, 即得到相应的R/T值, 该指标可反映某地植物区系的性质, 其值越大热带性质越强, 反之则温带性质越强[11, 23]。去除R/T为零的值后 (即不含热带分布型物种的样地) 对剩余的249个数据进行了对数转换, 使其接近正态分布以满足回归分析对因变量的正态分布要求后, 以海拔、年均温和年均降水量对其进行多元线性回归。
数据分析借助R软件 (Ver. 3.0.1, R Development Core Team, 2013) 完成, 其中NMDS排序、系统聚类以及MRPP检验均使用到vegan程序包[24]。地图绘制由ArcGIS9.3完成。
黄土高原山地天然或半天然森林群落的植物区系十分丰富, 仅就种子植物而言, 分别约占整个黄土高原种子植物区系科、属、种的66%、53%和37%[25]。根据野外样地调查结果, 黄土高原主要山地的天然、半天然森林群落由1222种维管植物组成 (含种下单元), 隶属于108科473属, 其中被子植物93科447属1179种 (含双子叶植物85科366属989种, 单子叶植物8科81属190种), 裸子植物4科7属9种, 蕨类植物11科19属34种。
本区森林中包含属较多的科 (≥ 10属):菊科 (Compositae, 46属)、禾本科 (Gramineae, 37属)、蔷薇科 (Rosaceae, 29属)、伞形科 (Umbelliferae, 22属)、豆科 (Leguminosae, 21属)、唇形科 (Labiatae, 19属)、毛茛科 (Ranunculaceae, 16属)、兰科 (Orchidaceae, 15属)、十字花科 (Cruciferae, 11属)、石竹科 (Caryophyllaceae, 11属); 含2~9属的科54个; 仅包含1属的科44个。包含种数较多的属 (≥ 10种):委陵菜属 (Potentilla, 22种)、蒿属 (Artemisia, 21种)、忍冬属 (Lonicera, 19种)、堇菜属 (Viola, 18种)、苔草属 (Carex, 17种)、风毛菊属 (Saussurea, 15种)、葱属 (Allium, 14种)、卫矛属 (Euonymus, 13种)、柳属 (Salix, 13种)、槭属 (Acer, 12种)、小檗属 (Berberis, 12种)、乌头属 (Aconitum, 11种)、黄耆属 (Astragalus, 11种)、拉拉藤属 (Galium, 11种)、蔷薇属 (Rosa, 11种)、悬钩子属 (Rubus, 11种)、绣线菊属 (Spiraea, 11种); 含有2~9种的属193个; 而仅包含1种的属249个。
依照吴征镒[2]的中国种子植物属分布区类型划分方案, 研究区内维管植物科的地理分布区类型构成有9种 (表2)。其中世界分布科最多 (达42个, 占总科数的38.9%, 依照惯例统计分析时未包括在内); 热带分布科共34个 (F2~F5、F7), 占统计分析总科数的51.5%, 温带分布科32个 (F8~F10、F14), 占统计分析总科数的48.5%。其中“ 泛热带分布” 型最多 (27科, 占40.9%), 其次是“ 北温带分布” 型 (23科, 占34.8%)。在科水平上, 热带分布与温带分布基本处于均衡状态。
科水平地理成分所对应群落尺度 (即每个样地) 的比例情况见图1A。由图易见, 群落尺度与前述 (以整个黄土高原为基准的) 区域尺度的统计结果总体上一致, 仍是世界分布科居多, 热带和温带分布科不分伯仲。两种尺度下的不同点在于群落尺度的“ 世界分布类型” 的比例更高 (平均为61.5%, 下同), 而“ 泛热带分布” (9.1%) 所占比例却普遍小于“ 北温带分布” (25.0%)。
研究区内维管植物属的地理成分共分为15个分布区类型 (表2)。其中世界分布属相对较少 (57个, 统计分析时未包括在内); 热带分布属 (F2~F7) 同样不多 (63个), 占总数的15.1%, 其主要类型为“ 泛热带分布” (31属, 占7.5%)。比较而言, 属级地理成分绝大部分具温带分布性质 (F8~F14, 共344属, 占82.7%), 其中主要类型为“ 北温带分布” (167属, 占40.1%), 其次为“ 旧世界温带分布” (67属, 占16.1%); 中国特有分布 (F15) 较少 (仅9属, 占2.2%)。属地理成分在群落尺度上的比例分布情况与区域尺度上的统计结果相类似 (图1B)。不同点在于, “ 北温带分布” 的地位更加凸显 (53.8%), 而作为热带分布主要类型的“ 泛热带分布” 在群落内所占比例很小 (3.0%)。
| 表2 黄土高原山地森林群落植物区系组成的科、属分布区类型 Tab. 2 Areal-types of the vascular plants in natural montane forests on the Loess Plateau |
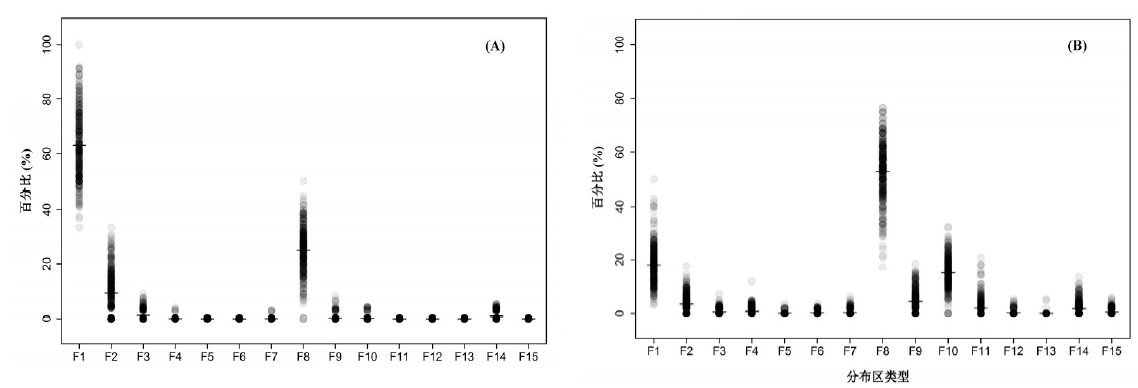 | 图1 群落尺度下黄土高原山地森林群落科 (A) 与属 (B) 的地理成分比例构成Fig. 1 Constituent ratio of area-types on the Loess Plateau counted by sampling plots at level of family (A) and at level of genus (B) |
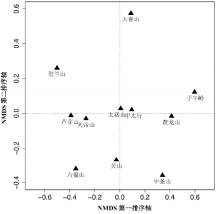
3.3.1 各山地间植物区系谱排序 从黄土高原各山地之间植物属一级区系谱矩阵的NMDS排序结果看 (图2)。排序的胁强系数 (Stress, S) 为0.052 (即5.2%), 属于拟合一般 (0.05 < S ≤ 0.10) 但接近良好的情形 (0.025 < S ≤ 0.05)[26]。
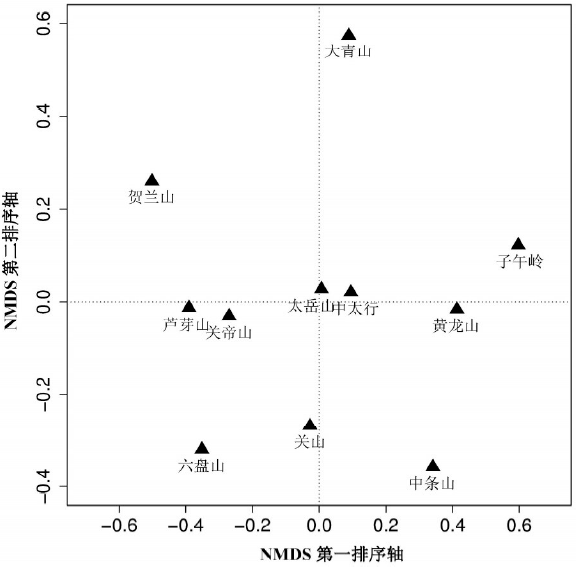 | 图2 基于属级“ 区系谱” 数据的黄土高原山地森林群落植物区系NMDS排序图Fig. 2 NMDS ordination diagram for 11 mountains on the Loess Plateau based on the matrix of genera floristic spectrum |
排序第一轴主要反映了各山垂直高度不同所引起的植物属垂直分异。沿该轴从左往右, 各山地海拔高度依次递减。以样地的平均海拔比较, 右边子午岭、黄龙山和中条山海拔最低 (平均1470~1630 m), 大青山、中太行 (左权) 和太岳山的海拔居中 (平均1670~1850 m), 最左面的五座山海拔较高, 一般在2000 m以上 (平均2010~2390 m)。排序第二轴则反映了山地的水平分布情况, 各山地沿该轴由北向南依次排开。属于同一山脉的不同山地在排序图上彼此位置接近, 表明其区系组成比例接近, 如同属吕梁山脉的芦芽山和关帝山, 中太行和太岳山, 同位于黄土高原中心位置的子午岭和黄龙山, 以及六盘山脉的六盘山与其南部支脉关山等。大青山和贺兰山则与其他山地相对距离较远, 反映出二者因居于区域北部和西北部边缘, 大陆性气候特征凸显, 对应在区系组成上与其他地区迥异。
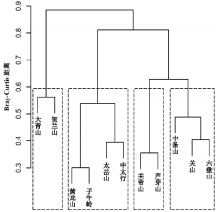
3.3.2 各山地间基于属水平的系统聚类 系统聚类在距离水平0.6时可将11座山地由北向南分为四组 (图3), 多响应置换过程 (MRPP) 检验表明, 四组之间差异显著 (9999次, p < 0.001)。第一组, 由西北部至北部的贺兰山和大青山组成, 并在系统聚类树中与其他山地分离较早。第二组由子午岭、黄龙山和太岳山、太行中部山地组成。第三组为吕梁山脉的芦芽山和关帝山。第四组由南部的六盘山、关山与中条山组成。比较聚类分析与NMDS排序结果, 两者呈现出基本一致的对应关系。
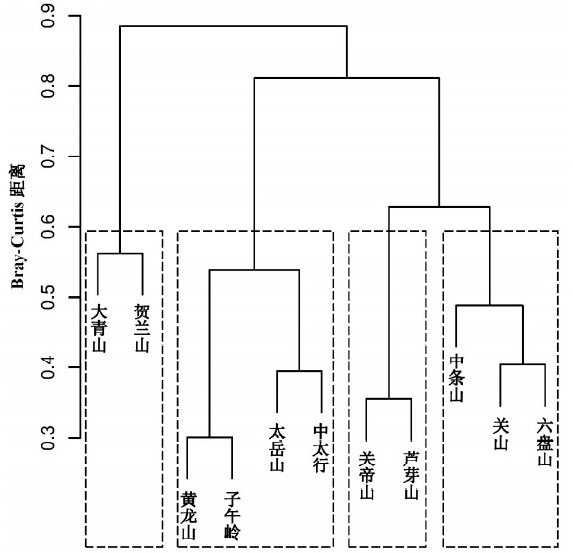 | 图3 黄土高原山地森林群落植物区系的聚类分析Fig. 3 Diagram of cluster analysis for 11 mountains on the Loess Plateau |
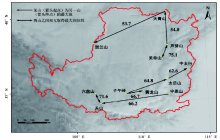
3.3.3 各山地间属的相似性 区系分析中一般认为, 当两地相似性系数低于50%时, 则表明二者间的区系联系较弱, 反之则较强[22]。不同山地两两间的Sø renson相似性系数变动在34.4%~75.1%之间。除大青山外, 其他山地与贺兰山的相似性均最低 (平均约36.6%), 其次是大青山 (平均约47.7%)。太岳山与其他各山地的平均相似性最大 (58.5%)将每座山地同与之相似性值最高的山地在地图上用双箭头连接起来 (图4), 可见同属一条山脉或空间上临近的山地往往相似性也最高。值得注意的是, 有两条箭头相连的山地 (如芦芽山、太岳山和关山等), 表明其和周围其他山地的相关度均较高, 也意味着该山地是物种迁移过程中的重要廊道。
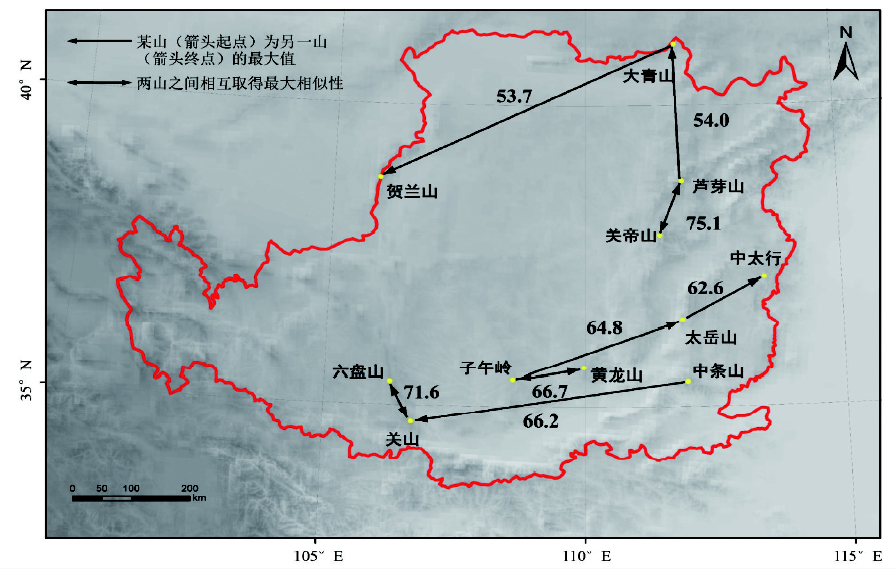 | 图4 黄土高原山地森林群落山地之间最大相似性系数Fig. 4 Maximum Sc indices between forest communities of 11 mountains on the Loess Plateau |
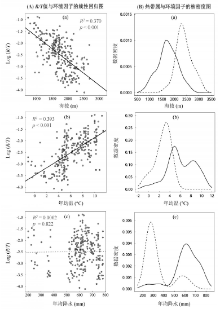
以海拔、温度和降水为自变量与样地R/T值 (对数) 作回归分析, 得到的多元线性模型能够解释数据42.1%的变异 (P < 0.001)。经比较标准化后自变量各自的偏回归系数 (表3), 发现年均温对R/T值的影响最大, 其次是海拔, 而年降水量则无明显影响。进一步分别对上述各自变量单独进行回归分析(图5)可看出, 群落R/T值随着海拔的升高呈下降趋势 (图5-A-a), 比较其对应热带属的多度分布图 (图5-B-a) 可知, 热带属的零值样地大多分布在较高海拔, 说明海拔高度是限制森林群落热带属植物分布的一个综合生态因子。R/T随着不同样地年均温的升高呈上升趋势 (图5-A-b), 而“ 零值点” 都分布于年均温小于5 ° C的样地内 (图5-B-b)。虽然R/T与年均降水量之间并无明显趋势 (图5-A-c), 但其“ 零值点” 则主要集中于280 mm左右, 而年降水小于450 mm的样地几乎没有热带属出现 (图5-B-c), 说明水分对该区热带属分布同样是重要的。
| 表3 多元回归模型的主要参数 Tab. 3 Main parameters of multiple regression model |
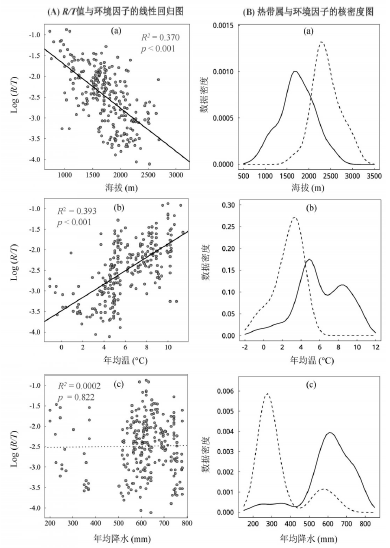 | 图5 黄土高原山地森林群落R/T值 (A) 与热带属 (B) 在生境梯度上的变化格局(图B中实线示热带属群落的分布情况, 虚线示无热带属群落的分布情况)Fig. 5 Patterns of the R/T values (A) and the Trop. Richness (B) within montane forest communities along with environmental gradients on the Loess Plateau |
本文发现, 在区域尺度上, 黄土高原山地森林群落内植物科、属的分布区类型, 均以温带成分 (特别是“ 北温带分布” 成分) 占主导地位, 这与其他学者的研究结果相一致[25, 27], 从而反映出黄土高原暖温带— 温带季风性半湿润至半干旱气候特点。在局地群落尺度上进一步研究发现, 温带分布型占主导的趋势依旧存在, 然而该类型所占比例在不同群落中往往存在较大差异 (图1)。这一现象一方面暗示了群落的物种构成是随机作用、环境筛滤以及自身生态位保守性等诸多因素综合作用的结果[28, 29], 不同质、量的因子组合将影响群落区系组成和结构上的差异。另一方面也说明, 研究对象在不同尺度内可能呈现出相异的规律, 区系地理研究同样应该重视生态学中普遍存在的尺度效应[30, 31]。由此可见, 区系属性可作为小尺度群落学研究中相对稳定的指标来厘定气候。
植物区系分析中, 植物属能够反映出植物系统发育过程的演化路径与地区性特征[32], 其分类学性状也相对稳定[23, 27, 33]。因此, 本文关于山地之间排序、系统聚类以及相似性计算等相关分析均基于“ 属” 级水平。我们发现, 起源相同、位置接近的山地, 其森林群落在属组成与分布区类型上往往表现出较高的相似性, 体现了地理要素在空间上的自相关性 (如关帝山与芦芽山之间, 子午岭与黄龙山之间等)。例外的情况则说明相似的群落由相似的局地环境决定, 空间自相关仅仅是群落相似性的必要条件。比如与中条山关系最为密切的并非空间距离与其最为接近的太岳山, 而是相对较远的关山, 二者有类似的纬度与海拔范围, 并且有秦岭作为两者间的“ 中介” 。
一般认为, 植被分布的地带性格局是水、热因子综合作用的直接结果[34]。通过回归分析我们发现, 年均温和海拔高度对研究区植物区系R/T值影响是重要的, 这两个因子都是反映某地热量条件的主要变量。热带属在黄土高原地区所呈现出的纬度递减格局和海拔递减规律, 也可间接支持多样性格局假说中的“ 寒冷忍耐假说” 相关论断, 即冬季低温是热带起源种北迁 (或垂直向上迁移)主要限制[35]。而水分条件的影响却并不显著, 其原因可能森林群落通常对水分的需求较高, 反过来能够支持高需水量乔木生长的环境, 其水分条件也比较优越。因此, 对于群落中其他生活型的物种来说, 水分并非其主要限制因子。对于R/T值为零的群落, 则很有可能代表了热带属在温带环境背景下对水热条件最低需求的上限, 亦即对应地理空间上的纬度北界或海拔上界 (图5-B-a、b)。曾有学者利用R/T值, 论证南方植物区系“ 平衡点” 问题[23, 36]。本研究结果显示, 虽然黄土高原属于远离该“ 平衡位置” 的地区, 但该值依然可作为定量探讨区系过渡性的优良指标。
对整个黄土高原范围内11座典型山地的353个样地数据统计分析, 获得以下结论:
(1) 黄土高原森林群落植物的分布区类型以“ 温带分布” 为主。黄土高原山地自然林群落野外调查发现维管植物共计108科473属1222种。基于区域和局地两种尺度的统计结果表明, 该区植物的科、属分布区类型以“ 温带分布” 特别是“ 北温带分布” 为主。
(2) 属的分布区类型与相似性充分展现出各山地之间的地理联系。以属为单元的有关分析发现, 山地之间植物区系联系能够较好反映它们在地理空间上的实际位置关系。从山地“ 区系谱” 数据矩阵出发, NMDS排序前两轴可以体现山地水平空间位置与海拔信息; 系统聚类分析可将研究区山地划分为4组, 该4组山地大体上具有由北向南的位置关系; 对山地间Sø renson相似性系数的量化分析, 得到的结果与排序、聚类相一致。
(3) R/T值能够反映植物区系的地带性格局。群落内R/T值以及热带属自身丰富度皆与研究区热量梯度密切相关, 具体呈现出纬度递减格局和海拔递减格局。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|